|
В. В. Бианки
ПРИРОДА КОЛЬСКО-БЕЛОМОРСКОГО РЕГИОНА
(краткое физико-географическое и биологическое описание)
К библиографии
Регион, где работают зоологи Кандалакшского заповедника, охватывает
весь Кольский п-ов с прилегающей к нему частью Баренцева моря и
все Белое море. Здесь на мореких островах, на акватории около них
и небольших участках побережья материка постоянно ведутся экологические
исследования, в которых основное место занимает изучение птиц. В
проводимой в этом направлении работе много внимания уделяется обыкновенной
гаге (Somateria mollissima), а также другим видам пластинчатоклювых,
ржанкообразных, хищных и воробьиных птиц. Работа проводится как
на стационарах заповедника, так и за его пределами, иногда на значительном
расстоянии. В 1958-1963 гг. при заповеднике находилась Северная
орнитологическая станция, где постоянно осуществлялось массовое
кольцевание птиц. Большую помощь сотрудникам заповедника оказывают
приезжие специалисты, студенты и специализированные группы юных
натуралистов.
Кольско-Беломорский регион расположен на границе двух разнородных
участков земной коры, в двух климатических и двух зоогеографических
областях.
Имеющиеся физико-географические описания региона не дают полного
представления о всем его разнообразии - они или слишком общие, или
их детали разбросаны по различным специальным изданиям. Предлагаемая
работа, надеюсь, даст достаточное представление о среде обитания
птиц и некоторых других животных, а также поможет поближе познакомиться
с ней биологам, которые только начинают работать в данном регионе.
Латинские названия сосудистых растений даются по сводке С. К. Черепанова
[101], моллюсков - по монографии "Моллюски Белого моря"
[63], других морских беспозвоночных - по последним работам сотрудников
Зоологического института АН СССР и Советскому энциклопедическому
словарю [87], рыб - по монографии А. П. Андрияшева [4], птиц - по
каталогу А. И. Иванова [43], млекопитающих - по сводкам В. Е. Соколова
[88].
Автор благодарен коллегам - ботанику Е. Г. Воробьевой, гидробиологам
Г. А. Шкляревич и Е. А. Нинбургу - за оказанную в подготовке материала
помощь.
В крайний Северо-Западный регион нашей страны
входит Кольский п-ов с прилегающей к нему с запада частью материка
и прибрежной полосой шельфа Баренцева моря, а также Белое море с
прилегающими к нему Северной Карелией и северо-западной частью Архангельской
области, включая п-ов Канин. Регион расположен на стыке двух крупных
участков земной коры - Балтийского кристаллического щита и Русской
платформенной равнины, которые имеют значительные различия в геологической
структуре, строении и истории. На Кольском полуострове и в Карелии
архейские породы Балтийского щита - граниты, гнейсы, диабазы - прикрыты
тонким слоем осадочных пород и нередко выходят на дневную поверхность.
С нижнепалеозойского времени щит не покрывался морем, здесь преобладали
процессы разрушения. Мощные ледники выпахали осадочные породы, сгладили
возвышающиеся обнажения материнских пород и оставили массу моренного
материала. Наибольшие высоты рельефа отмечаются на северо-западе
региона. Это Хибины - до 1200 м н.у.м., Ловозерские тундры и Чуна-тундра
- до 1120 м, , Сальные тундры - до 1000 м н. у. м. и др. Близ Кандалакши
высота гор меньше - до 780 м, а южнее, у оз. Топозера в Северной
Карелии, - до 580 м н.у.м.
Саами (лопари) называли возвышенности, на которых отсутствовал лес,
тундрами, а покрытые тайгой небольшие скалистые гряды - вараками.
Было предложено сохранить название "вараки" за возвышенностями
до 200 м, более высокие, от 200 до 400 м, называть "тунтури"
(от финского "tunturi" - гора, сопка), а выше 400 м -
горами или, как принято в Прибеломорье, тундрами, поскольку этот
термин сохраняется в географических названиях [48].
О. Н. Казакова различает в Мурманской области 27 ландшафтов, из
которых 16 относит к трем округам Таежной провинции, 4 - к одному
округу Лесотундровой провинции и 7 - к двум округам Тундровой провинции.
Наиболее разнообразны Кольский горный округ (8 ландшафтов), Варзуго-Терский
(6 ландшафтов) и округ примореких тундр, состоящий из 5 хорошо различающихся
ландшафтов [48].
У основания Кольского п-ова горные системы разной величины и строения
чередуются с депрессиями. Пологие склоны местами сменяются крутыми
обрывами, часто встречаются глубокие ущелья. Значительная пересеченность
местности создает большую мозаичность экологических условий и размещения
животных и растений, что в свою очередь способствует поддержанию
благоприятных условий для проникновения видов в соседние ценозы.
Западнее 36° в. д. земная кора поднимается. Наиболее быстрый подъем
отмечается близ кута Кандалакшского залива - около полуметра в столетие
[70]. В результате море отступает, образуются новые участки суши,
острова.
Восточное побережье Белого моря является западной окраиной Русской
платформы. В палеозойское и мезозойское время эта территория неоднократно
покрывалась морем. Ее кристаллический фундамент накрыт пластом мягких
осадочных пород толщиной 500-1000 м у Зимнего берега и до 4000-5000
м у западных склонов Тимана. У южных и восточных берегов Белого
моря находятся лишь небольшие возвышенности - сильно размытый древний
кряж Ветреный Пояс (до 200-350 м н. у. м.), расчлененный на отдельные
вершины (Синие горы), которые поднимаются над заболоченной равниной
на 60-80 м. Остальная часть территории входит в Онего-Двинско-Мезенскую
равнину, имеющую слабый уклон к северу. Водоразделы здесь плоские,
заболоченные, иногда холмистые, не превышающие 300 м н. у. м. На
северо-востоке Прибеломорья Канинская тундра не поднимается выше
100-150 м н. у. м. за исключением платообразной возвышенности Канин
Камень (Поэ) на самом севере, достигающей 240 м [49, 81].
В противоположность Северной Карелии и основанию Кольского п-ова
восточное побережье Белого моря опускается примерно с той же скоростью,
с какой поднимается на западе. Море наступает на свои восточные
берега, подмывая и разрушая их. Сильное влияние на формирование
современных ландшафтов оказали здесь потепление и похолодание климата,
происходившие после ледникового периода.
Рассматриваемый регион находится между 64° и 70° с. ш., через его
середину проходит Северный полярный круг. На юге региона в летнее
время в течение почти двух месяцев светлые сумерки заменяют темные
ночи и день составляет более 19 часов. На самом северном полуострове
- Рыбачьем - светлые ночные сумерки (так называемые гражданские,
когда солнце находится за горизонтом под углом не более 6°) начинаются
25 апреля и заканчиваются 15 августа. С 20 мая по 25 июля солнце
здесь совсем не опускается за горизонт. В вершине Кандалакшского
залива гражданские сумерки начинаются 5 мая и продолжаются до 5
августа. Кроме того, выделяют еще навигационные сумерки, когда солнце
находится под углом 6-12° по отношению к горизонту. В Кандалакше
они начинаются 20 апреля и кончаются 25 августа (рис. 1).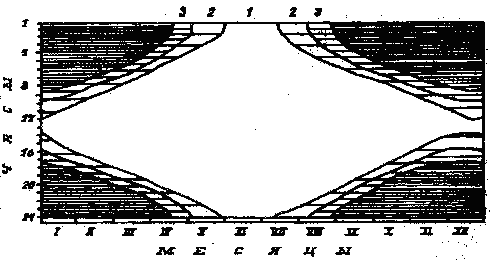
Рис. 1. Продолжительность светлого времени суток на широте г. Кандалакши:
/ - солнце над горизонтом; 2 - гражданские сумерки;
3 o навигационные сумерки
Регион находится под влиянием арктического и умеренного морского
воздуха Атлантики, приносящего с собой муссонные ветры. Климат региона
является переходным от морекого к континентальному с часто повторяющимися
циклонами, которые зимой сопровождаются оттепелями [16]. По классификации
климатов [2] регион занимает две области двух поясов: Атлантическую
область Субарктического пояса и Атлантико-арктическую область Умеренного
пояса. Первая из них охватывает южную часть Баренцева моря, тундровую
полосу Мурманского побережья и п-ов Канин. Она характеризуется как
влажная, умеренно холодная. Суммарная солнечная радиация составляет
здесь 65 ккал/см2 в год, сумма температур воздуха выше 10°-200°.
Вторая область охватывает расположенную южнее лесную часть региона.
Характеризуется как избыточно влажная и умеренно теплая: суммарная
солнечная радиация составляет здесь 75-80 ккал/см2 в год, сумма
температур воздуха выше 10° - 800°-1200°. Белым морем и р. Онегой
область делится на две подобласти - восточную и западную.
Климат Мурмана формируется непосредственно под влиянием теплого
течения Гольфстрим, часть которого (Нордкапская ветвь) омывает Западный
Мурман. Море и глубоко вдающиеся в материк узкие заливы здесь никогда
не замерзают. Вдоль Восточного Мурмана проходит ослабленная прибрежная
ветвь Мурманского течения, которое является продолжением части Нордкапской
ветви. И хотя обычно море здесь тоже не замерзает, в суровые зимы
лед может распространиться с востока до о-ва Кильдин и покрыть заливы.
Степень ледовитости Восточно-Мурманского прибрежья Баренцева моря,
кроме температуры, зависит от тепловой мощности Нордкапского течения
и ветрового дрейфа льдов. В колебаниях ледовитости прослеживается
11-13-и, по-видимому, 25 - 27-летняя периодичность [93]. С появлением
льда на острова по ледяным мостам переходят лисицы (Vulpes vulpes)
и белые медведи. Оставаясь там годами, они уничтожают доступные
им гнезда, птенцов и взрослых птиц. :
На Мурмане за год бывает до 80-120 штормовых дней, большая часть
которых приходится на зиму. Ветры сдувают снег с возвышенностей,
нередко оголяя их, и наметают сугробы в понижениях рельефа. Зимой
тут тепло, летом прохладно, средняя месячная температура воздуха
составляет зимой -10°, летом +10°. В течение всего года отмечаются
резкие перепады температуры. С продвижением на восток климат становится
более суровым, а с удалением от побережья Баренцева моря - более
континентальным (табл. 1).
| Районы |
|
Безморозный период |
т. Сумма температур и продолжительность |
т. Сумма температур и продолжительность |
т. Сумма температур и продолжительность |
т. Сумма температур и продолжительность |
Сумма осадков, мм |
Сумма осадков, мм |
|
местонахождение |
|
5 гр |
5 гр |
10 гр |
10 гр |
за год |
в летнее время |
| 1 |
Горные арктические пустыни, полоса
побережья от Святоносского залива к Лумбовскому и
далее к заливу Панфилова
|
90-100 |
- |
90-100 |
300 |
0-160 |
300-380 |
- |
| 2 |
П-ов Рыбачий с участком материка,
полоса побережья от Кольского залива до о-ва Сосновец |
100-105 |
880-1000 |
85-105 |
300-500 |
15-25 |
300-433 |
- |
| 3 |
Лесотундра, береговые криволесья
тайги |
90-100 |
1040-1170 |
95-110 |
500-700 |
40-50 |
300-400 |
190-240 |
| 4 |
Северо-восточный участок тайги,
окаймляющий горные поднятия в материковой части |
60-80 |
1180-1300 |
110-115 |
700-900 |
50-60 |
350-640 |
200-340 |
| 5 |
От оз. Имандра до р. Варзуги и
берега Белого моря, а также к Ковдору |
80-100 |
1300-1400 |
120 |
900-1000 |
60-70 |
340-570 |
220-270 |
| 6 |
От г. Кандалакши до Карелии и
Алакуртти; от Канозера расходящимся углом к Белому морю |
80-100 |
1400-1500 |
120-125 |
1000-1200 |
70 |
360-500 |
200-310 |
Таблица I
Агроклиматические районы Мурманской области
Примечание. Характеристика районов дана по Агроклиматическому справочнику,
1961Климатообразующими факторами Белого моря, кроме солнечной радиации
и атмосферной циркуляции, являются речной сток и водообмен с Баренцевым
морем [42]. Для Терского берега Горла Белого моря характерны резкие
гидрометеорологические аномалии, связанные с интенсивным перемешиванием
приливно-отливными течениями небольшой в этом месте толщи воды,
такие, как потеря теплой струи Мурманского прибрежного течения,
запаздывание фаз годового хода и уменьшение колебания температуры
воздуха и воды, а также другие аномалии [6].
Зимой нередки вторжения воздушных масс с Атлантического океана,
сопровождающиеся сплошной низкой облачностью и ослаблением морозов.
Приходящий из Арктики антициклон влечет за собой северо-восточные
ветры и довольно сильные морозы (ниже -20°). Весной погода неустойчива,
направление местных ветров часто зависит от конфигурации берега,
нередки туманы [37]. Лето прохладное и умеренно влажное. На острова
осадков выпадает меньше, чем на прибрежные возвышенности. Температура
воздуха держится в пределах 16-20°, в отдельных случаях поднимаясь
до 30°. На осеннее похолодание значительное влияние оказывает температура
водных масс, нивелируя температурные различия внутриматериковых
частей Белого моря. Часты штормы.
Зимой море покрывается льдом - полосой берегового припая разной
ширины и дрейфующими льдами. В заветренных опресненных заливах лед
появляется уже в октябре, а большая часть открытой акватории замерзает
обычно в декабре. В шхерных районах лед местами подмывается снизу
приливно-отливными течениями, кое-где встречаются полыньи. Последние
наблюдаются и во многих местах у границы берегового припая.
Вскрытие Белого моря происходит в мае, хотя рост полыней и появление
новых наблюдается в апреле, а иногда и в марте. Часть льда тает
на месте (особенно на литорали), значительное количество его выносится
постоянным течением вдоль Зимнего и oКанинского берегов в Баренцево
море.
Климат Северной Карелии более однороден, чем климат Кольского п-ова,
и, несмотря на ее значительную широтную протяженность, малоизменчив.
Сильное влияние на него оказывают близость Белого моря и повышающийся
к западу и северо-западу рельеф. В Прибеломорекой низменности весенние
заморозки кончаются, а осенние начинаются на 1,0-1,5 недели позже,
чем в приозерноравнинном Калевальском районе. Безморозный период
близ Белого моря продолжается в среднем 120 дней, в более отдаленных
местах - 181 день. В летнее время сумма дней с температурой воздуха
выше 10° составляет у побережья 70 дней, в западной части - 85 дней.
Средняя дата наступления первого мороза приходится у побережья на
20 сентября, а зима начинается здесь во второй половине октября
[82].
Климат восточной подпровинции более континентален, чем западной,
меньше подвержен изменчивому влиянию Гольфстрима, годовые колебания
его незначительны. Зимой в 50% случаев погода стоит "значительно
морозная" и "сильно морозная", 2% приходится на "жестоко
морозную", снег держится 185-200 дней. Летом сумма положительных
температур больше 10° составляет 900-1250°. На западе преобладает
в основном "умеренно морозная" и "значительно морозная"
зима, снег лежит 180-220 дней. Сумма положительных температур воздуха
больше 10° составляет 600-1200° [81].
Реки Карело-Кольского края имеют сравнительно небольшие бассейны
и отличаются значительной озерностью. Наибольшая водосборная площадь
отмечается у рек Кемь, Выг и Ковда - по 26-28 тыс. кв. км, а на
Кольском п-ове - у Поноя и Нивы - по 13-15 тыс. кв. км. С. В. Григорьев
[30] разделил реки по местонахождению в их бассейнах озер на 5 типов:
каскадные - со ступенчатым профилем и более или менее равномерным
расположением озер - Умба, Ковда (до строительства каскада ГЭС);
с верховым регулированием - озера сосредоточены в верховьях рек
- Кола, Воронья, Кемь;
сбросовые - небольшой длины водотоки, часто с большим уклоном: Нива,
Колвица;
безозерные - собирающие воды прямо из болот - Поной и др.;
смешанные - относящиеся в разных частях своего течения к различным
типам - Кереть и некоторые другие.
Ледостав на реках приходится в среднем на конец октября -середину
ноября. Многие пороги не замерзают, и на них могут зимовать водоплавающие
птицы. Вскрытие рек происходит в начале мая, а полыньи на них появляются
в апреле. Паводок нередко начинается еще при ледоставе в середине
мая, а на мелких реках - не выражен совсем. Наибольший уровень воды
отмечается в конце мая-июне, межень - в июле-августе. Дождевые летние
паводки незначительны, хотя в местах с малым уклоном они могут оказывать
неблагоприятное влияние на гнездование птиц, селящихся на их берегах.
Большинство рек Кольского п-ова и Северной Карелии относится преимущественно
к снежному типу питания, меньшее число - к дождевому типу питания
и совсем мало - к подземному. Мутность воды обычно слабая - меньше
20-25 г/м3, минерализация также незначительная - меньше 100 мг/л.
Реки Кандалакшского, Карельского и Поморекого берегов, на территории
питания которых преобладают граниты и гнейсы, имеют минерализацию
до 25 мг/л. При наличии легкорастворимых пород - сланцев и карбонатных
соединений - в бассейнах рек Терского берега и девонских образований
на Лямицком берегу средняя минерализация повышается до 40 мг/л и
более. Наибольшей минерализацией обладает вода р. Онеги - более
100 мг/л. Большинство рек относят к гидрокарбонатному типу, обычно
к группе кальция, реже магния [62]. Количество растворенного в них
гумуса прямо пропорционально заболоченности водосбора и обратно
- его озерности [31].
Характеризуя растительность рек северо-востока Кольского п-ова,
Ю. Д. Цинзерлинг и Е. К. Косинская (1935) пишут, что в ручьях и
быстротекущих речках высшая растительность отсутствует, а в реках
она очень незначительна. Исключение составляет только р. Поной.
Главные реки Русской равнины, впадающие в Белое море, - Онега, Северная
Двина, Мезень - имеют хорошо разработанные долины и широкие поймы.
Водоразделы здесь часто заболочены, а между Пинегой и Кулоем во
время весеннего половодья они сливаются. При малом уклоне скорость
течения рек незначительна и высокий уровень воды нередко сохраняется
длительное время. Однако следует отметить, что "пограничная"
между западом и востоком р. Онега порожиста, а р. Емца (левый приток
Северной Двины) в верховьях зимой не замерзает вследствие подземно-карстового
питания и быстрого течения на порогах [22].
Самая большая река Белого моря - Северная Двина. Площадь ее бассейна
- 360 тыс. кв. км, годовой сток - 110 млрд. куб. м воды [22]. Несколько
ниже устья Пинеги Северная Двина разбивается на многочисленные протоки
с островами между ними. Широкая долина, достигающая местами 10 км,
у Холмогор становится еще шире - до 18 км. Ниже Архангельска на
протяжении 45 км река образует обширную дельту площадью 11 тыс.
кв. км [35].
Вторая по величине р. Мезень имеет площадь водосбора 76,5 тыс. кв.
км. Нижний участок реки на протяжении более 60 км испытывает влияние
морекого прилива, что характерно также для Северной Двины и других
рек Русской равнины, впадающих в Белое море. Площадь бассейна р.
Онеги составляет '57,5 тыс. кв. км, остальных рек - менее 5 тыс.
кв. км.
Освобождение рек Русской платформы ото льда происходит во второй
половине апреля - первой декаде мая, замерзание - в ноябре. Мутность
и минерализация воды хотя и больше, чем в Карело-Кольском крае,
но все же довольно слабые - обычно меньше 50 г/м3 и 200 мг/л соответственно.
Большинство рек также гидрокарбонатного класса. Вода многих из них,
особенно крупных, в последнее время сильно загрязнена различными
отходами производства.
Карелия и Кольский п-ов являются одним из самых озерных мест нашей
страны. В Карелии насчитывают 30153 озера площадью от 1 га и больше,
которые принадлежат бассейну Белого моря. Площадь их зеркала составляет
11043,4 кв. км. Озер с площадью зеркала менее 1 кв. км насчитывается
29308, а меньше 10 га - 24436; суммарная площадь их зеркала составляет
2018,5 и 682,3 кв. км соответственно [30].
Наибольшее количество озер в Карелии расположено между 65°30"
и 66°00" с. ш., а к северу, востоку и югу число их уменьшается,
особенно за 35° в. д. [29]. В Мурманской области находится 11600
озер общей площадью 9200 кв. км. Большинство их расположено в восточной
части Кольского п-ова. Вскрытие мелких озер отмечается в середине
мая, крупных - позже, иногда только в середине июня. Осенью мелкие
озера замерзают во второй половине октября, крупные - в ноябре.
Озера Северной Карелии и Кольского п-ова содержат низко минерализованные
воды подзолисто-болотных почв с незначительным количеством биогенных
веществ и, следовательно, имеют низкую биологическую продуктивность.
В бассейне Белого моря расположены озера, которые относят преимущественно,
к гумифицированной группе (со значительным поступлением в них болотных
вод). Гумификация способствует переходу некоторых биогенных веществ
в коллоидное трудноусвояемое состояние, в результате чего понижается
их продуктивность [7б]. Прозрачность воды в озерах слабая, обычно
не более 3 м, только в олигогумозных (Топозеро, Пяозеро и др.) она
увеличивается до 6 м.
Характеризуя растительность озер северо-востока Кольского п-ова,
Ю. Д. Цинзерлинг и Е. К. Косинская делят их на 3 группы [99]. Наиболее
многочисленную группу составляют озера с каменистым или песчано-каменистым
дном, бедным составом водорослей. Высшая растительность в таких
озерах отсутствует совсем или образует вдоль берега кайму из осок
(р. Сагех). Во вторую группу входит небольшое число озер с торфянистым
дном, берега которых обычно окаймлены зарослями осок. И третья группа
- это озера с глинистым дном, богатые водной растительностью.
Исследование оз. Вялозеро (водное зеркало - 103 кв. км, площадь
водосбора - около 450 кв. км) показало, что его растительный покров
однообразен и представлен 19 ассоциациями. В прибрежной растительности
наиболее распространены фитоценозы осок (Сагех sp.) и вахты (Menyanthes
trifoliata), хвоща (Equisetum sp.) и тростника (Phragmites communis),
а также другие виды растений. В июле-августе 1968 г. биомасса зоопланктона
колебалась от 1,8 до 28,8 г/м2, а бентоса в разных частях озера
- от менее 1 г/м2 до 10 г/м2. Большую роль в населении дна играют
личинки хирономид [23].,
Количество зоопланктона в малых озерах довольно различно. В олиготрофных
оно обычно не превышает 1,0-1,5 г/м3, в мезотрофных - 2,0-3,0, а
в эвтрофных достигает 5-7 г/м3 и более. Биомасса планктона в значительной
степени зависит от рН среды. В олиготрофных малокислотных озерах
число видов рачков и коловраток почти на 50 % меньше, чем в озерах
с нейтрально-щелочной водой, а в среднекислотных - меньше на 60-65
%. В полиацидных (кислотных) водоемах число форм планктона более
чем в 3 раза уступает их численности в нейтрально-щелочных озерах
[84].
Молодь большинства промысловых рыб кормится бентосом. Более чем
в половине озер его биомасса низкая - в среднем менее 5 кг/га (табл.
2), в некоторых крупных озерах (Энгозеро, Тикшозеро, Керетьозеро
и др.) на мелководьях и в отдельных губах, а также в части небольших
озер - до 10 кг/га. Значительную долю биомассы бентоса составляют
личинки хирономид (Chironomidae), олигохеты (Oligochaeta) и другие
беспозвоночные. По преобладающей группе беспозвоночных, которыми
питаются рыбы, С. В. Герд [25] выделил 3 типа озер:
тендипедидный, олигохетный (наиболее кормный) и амфиподный, а по
месту обитания рыб - 4 зоны: верхняя литораль (до 2-3 м глубины),
нижняя литораль (до 6-8 м), зона средней глубины (до 20-25 м) и
зона большой глубины (свыше 25-30 м). На Кольском п-ове большинство
озер тундры, как и реки, бедны растительностью и гидробионтами,
вследствие чего заселяются преимущественно прибрежными птицами,
которые кормятся на берегу, - гуменниками (Anser fabalis), куликами
(Limicolae) и др.
Л. А. Гибет разделила озера Карелии как охотничьи угодья на 4 типа:
травянистые, смешанные, каменистые и черные ламбины [26]. Два первых
типа интенсивно используются водоплавающими птицами: травянистые
- гусями, речными утками и хохлатой чернетью, смешанные - гагарами,
лебедями, гоголями и крохалями. На каменистых озерах и черных ламбинах
птиц встречается мало.
| Природные зоны |
Районы |
Средняя первичная продукция |
Биомасса г\м |
|
|
|
|
|
Зоопланктон |
Бентос |
Зоопланктон + бентос |
| Тундра |
Кольский |
0.22 |
3.06 |
2.27 |
5.61 |
|
Большеземельский |
0.35 |
2.68 |
6.06 |
9.01 |
|
В среднем |
0.24 |
2.74 |
4.64 |
7.86 |
| Тайга |
Мурманская область |
- |
2.99 |
2.60 |
5.42 |
|
Северная Карелия |
- |
2.32 |
4.16 |
6.71 |
|
Архангельская область |
- |
4.01 |
3.05 |
7.64 |
|
в среднем |
1.56 |
4.51 |
3.90 |
9.55 |
Таблица 2
Продуктивность озер тундры и северной тайги в Кольско-Беломорском
регионе (С. П. Китаев, 1984)
В малых озерах о-ва Великого в Кандалакшском заливе
-ламбинах, окруженных болотной сплавиной, с низкой кислотностью
воды и в глубоких проточных озерах с холодной водой, имеющих слабощелочную
реакцию, средняя биомасса планктона составляет менее 1 г/м3, бентоса
- менее 3 г/м2. Птицами они практически не заселяются. Зато сточные
озера со слабым притоком болотной воды и слабокислой реакцией, прозрачностью
1,0-1,7 м, имеющие среднюю биомассу планктона - 9-17 г/м3 и бентоса
- 2,2-11,5 г/м2, заселены речными утками (р. Anas), хохлатыми чернетями
(Aythya fuligula) и серощекими поганками (Podiceps griseigena).
Сходная биомасса бентоса была обнаружена в оз. Мураканском на Летнем
берегу Онежского п-ова. При площади водного зеркала в 1,5 тью. га
и наибольшей глубине 5 м оно имеет разнообразное население дна со
средней биомассой 2,5 г/м2. Преобладающую ее часть составляют личинки
хирономид - до 2,2 г/м2, моллюски, главным образом катушки (Planorbidae)
и горошинки (Pisidiidae), - до 1,25 г/м2, ручейники (Trichoptera)
-до 0,8 г/м2. Наибольшая биомасса - до 8,5 г/м2 - отмечена на илистом
грунте открытой части озера, меньшая - на песках - до 6,8 г/м2 -
в северной части и 6,2 г/м2 - на литорали [71].
Многие крупные озера региона, такие, как Имандра, Умбозеро, Ловозеро,
Пяозеро и ряд других, используются как водохранилища или для сброса
промышленных и бытовых сточных вод. В результате их естественная
гидрохимическая и гидробиологическая характеристики сильно изменены,
а продуктивность снижена. Крупные озера сейчас мало используются
птицами. Зато те немногие из них, которые имеют богатую водную растительность
и повышенную биопродуктивность, заселяются различными видами водоплавающих
и околоводных птиц.
В Архангельской области находится около 2,5 тысяч озер, большинство
из которых располагается в aa западной части, примыкающей к Карелии,
а также в поймах крупных рек [17]. Общая площадь озер составляет
более 5 тыс. кв. км, а небольших озер площадью до 1 кв. км, которых
насчитывается 64%, - около 410 кв. км [35].
Гидробиологического описания озер восточного побережья Белого моря
нам не удалось найти. Пойменные озера, вероятно, достаточно богаты
и вполне подходят для гнездования на них и остановки во время миграции
многих водоплавающих и прибрежных птиц. Примером может служить Пинежский
заповедник на правобережье р. Пинеги, на территории которого находится
83 озера. Птицами там заселены преимущественно крупные озера, расположенные
на моренной равнине и имеющие богатую растительность. Мелкие озера
площадью 1-3 га обычно карстового происхождения и бедны растительностью.
Учет, проведенный в мае-июне 1977-1982 гг., показал, что на 141-227
га водной поверхности встречено в среднем по 78 водоплавающих птиц
на 10 га. Наиболее многочисленными из них были большой крохаль (Mergus
merganser) - по 21 экз., хохлатая чернеть - по 16 экз. и гоголь
(Bucephala clangula) - по 15 экз. Меньше было гуменников (Anser
fabalis) - по 10 экз., крякв (Anas platyrhynchos) - по 8 экз. и
чернозобых гагар (Gavia arctica) - по 5 экз. Остальные виды представлены
незначительным числом особей [83].
С учетом особенностей рельефа и климата растительность Кольско-Беломорекого
региона относят к подзонам южной кустарниковой подпровинции Циркумполярной
тундровой области северной тайги Кольско-Печорской и Валдайско-Онежской
подпровинции Евразиатской таежной области [47]. Тундры и предтундровые
редколесья в сочетании с болотами тянутся вдоль Мурманского побережья
широкой полосой от 75 до 100 км, занимают весь Канин п-ов и переходят
в узкие полосы вдоль Зимнего берега Горла Белого моря и Абрамовского
берега Мезенского залива (рис. 2). Кроме того, ими покрыты расположенные
южнее горные возвышенности.
Кустарниковые тундры простираются вдоль побережья Баренцева моря
полосой, ширина которой изменяется от 20 км на западе до 120 км
в бассейне р. Иоканги. Далее на восток полоса тундр сужается и по
Терскому берегу доходит до устья р. Стрельны [100]. На п-ове Рыбачьем
и на Восточном Мурмане у берега находятся щебнисто-каменистые тундры
и скальные обнажения, поверхность которых покрыта растительностью
только на 20-40%. Дальше от моря лежит полоса кустарничковых и ерниковых
тундр. Первые отличаются обилием в них бореальных видов - черники
(Vaccinium myrtillus), брусники (V. vitis-idaea), хвощей (Equisetum
pratense, E. sylvaticum), луговика-щучки (Lerchenfeldia flexuosa).
Вороника (Empetrum hermaphroditum) со шведским дереном (Chamaepericlymenum
suecica) местами образуют специфический для этого региона тип тундр
[28]. В ерниковых (кустарниковых) тундрах преобладает примесь сизой
ивы (Salix glauca), иногда сибирского можжевельника (Juniperus sibirica),
а в кустарничковых, кроме уже перечисленных, -примесь голубики (V.
uliginosum), вереска (Calluna vulgaris), овечьей овсяницы (Festuca
ovina), осок (Carex brunnescens, С. bigelowii) и других растений.
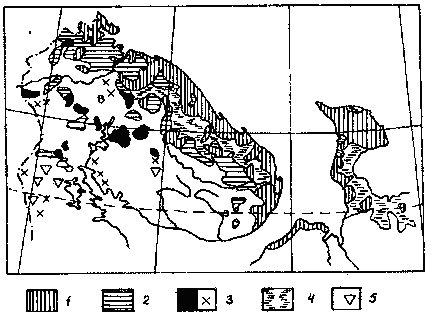
Рис. 2. Расположение тундровых ландшафтов, болот
и редколесья
(по С. А. Грибовой, 1980);
1 - южные тундры; 2 - предтундровые березовые редколесья;
3 - горные тундры; 4 o болота; S - горные редколесья ,
Ивняковые тундры встречаются преимущественно на северо-востоке.
Крупноерниковые травяно-кустарничковые зеленомошные тундры располагаются
обычно у подножия склонов. Они имеют 3 хорошо выраженных растительных
яруса: сильно сомкнутые кустарники высотой 50-100 см, густой травяно-кустарничковый
покров высотой 10-15 см и сплошной моховой ковер из Pleurozium schreberi
с примесью других видов и отдельных кустиков лишайников р. Cladonia
[28]. Широко распространены также березовые редколесья и криволесья
[80]. В бассейнах рек Вороньей, Харловки, Иоканги они длинными "языками"
проникают на север.
Господствующая на больших площадях во всех типах тундр вороника
является основной пищей для птиц открытых пространств - гуменников,
средних кроншнепов (Numenius phaeopus), чаек (р. Larus) и других.
На островах Кандалакшского залива вороника дает в пересчете на 1
га 0,5 т ягод при урожайности 3 балла по шкале Каппера-Формозова
и более 2,0 т при урожайности в 5 баллов [15]. Своеобразную помощь
оказывают птицам пасущиеся здесь стада северных оленей (Rangifer
tarandus). Добывая зимой корм, они раскапывают снег, который весной
в таких местах быстрее стаивает, и на обнажающейся земле кормятся
прилетевшие птицы.
Обширные площади в тундре и лесотундре занимают болота. Большей
частью это травяно-лишайниково-моховые бугристые болота, меньше
- травяно-сфагново-гипновых типа аапа [98, 18]. Для первых характерно
наличие осоки (Carex sp.), пушицы (Eriophorum sp.), а также ерников
и кустарничков, для вторых -деревьев, кустарников и разнообразных
болотных, водно-болотных и водных трав [103]. По окраинам болот
и в местах с увлажненными почвами растут ивняки (Salix sp.), образуя
то разреженные, то труднопроходимые заросли высотой до 50 см 100],
где гнездятся некоторые воробьиные птицы (Passeriformes).
Южнее озерно-болотной полосы наблюдается повышение уровня местности,
некоторые вершины достигают почти 400 м н. у. м. Это Кейвы. В их
западной части находятся глубокие ущелья с отвесными скалами, в
восточной - пологие плакорные участки. Здесь широко представлены
массивы крупноерниковых кустарничковых лишайниковых тундр, понижения
заняты участками предтундровых лесов.
Особое место на северо-западе региона занимают горные тундры, лежащие
на склонах и вершинах гор выше 350-500 м I. у. м. Выше 750-1000
м на Хибинах, в Ловозерских горах, на Чуна- и Монче-тундрах расположены
высокогорные арктические пустыни. В горных массивах, островами возвышающихся
среди северной тайги вплоть до оз. Пяозера, проходит пояс субальпийского
криволесья, над ним - кустарниково-лишайниковые тундры. Еще выше
находятся арктические пустыни, мохово-лишайниковая растительность
которых покрывает менее 5 % их площади. Зимой ветры сдувают снег
в понижения, где он местами не успевает растаять до следующей зимы.
Вегетационный период длится там всего 35-50 дней [100].
На п-ове Канин большие площади покрыты заболоченными мелкоерниковыми
тундрами, которые мозаично перемежаются с бугристыми болотами. Характерными
элементами тундр являются крупно-ивняковые участки с хорошо развитым
травяно-кустарничковым ярусом и зарослями нескольких видов ив высотой
до 1,5 м и с большой сомкнутостью крон (60-80 %). На перешейке между
реками Шойной и Мглой среди сплошных массивов бугристых и грядово-мочажинных
болот типа аапа отдельными островками расположены крупноерниковые
заболоченные тундры, которые встречаются и на южном берегу Мезенской
губы [28].
Всю остальную территорию региона южнее лесотундры занимает североевропейская
таежная провинция, которая западнее Белого моря делится на 2 широтные
полосы -редкостойных и осветленных лесов [80]. К первой относят
южную часть Мурманской области (кроме Ковдозерской низины) и северо-западную
часть Карелии. Здесь господствуют разреженные сосняки и ельники
высотой до 14-16 м. Сосняки больше приурочены к западной части территории,
ельники - к восточной и юго-восточной. В обоих типах леса много
березы (Betula sp.).
Особым флористическим богатством выделяются Хибинские и Ловозерские
горы, являющиеся восточным форпостом западной, гористой части Мурманской
области. Пограничная линия между низменной, сильно заболоченной
восточной частью и гористой, пересеченной западной проходит через
озера Ловозеро, Умбозеро, юго-восточную часть Имандры и Колвицкое.
К Кандалакшскому заливу она выходит между Колвицкой и Порьей губами.
В. Ф. Цветков и В. Г. Чертовской [97] делят Мурманскую область на
16 лесорастительных районов (табл. 3, рис. 3). Наиболее лесные из
них находятся в южных и западных частях области, а заболоченные
- в юго-восточных. В половине земель всех выделенных районов воды
занимают по 10-26 %, а в шести из них по 13-52 % приходится на тундры.
В древостое сосны (Pinus sp.), как правило, преобладают над елями
(Picea sp.), лишь в двух районах преобладающей породой является
береза. Средняя сомкнутость крон составляет: сосняков - 0,39-0,48
(в Ковдозерском районе - 0,52), ельников - 0,35-0,44, березняков
-0,41-0,49.
| Категории земель, % от общей площади |
|
|
|
|
Средневзвешенный состав пород |
| Районы |
Леса |
Воды |
Болота |
Гольцы и тундры |
|
| Печенгский |
48,6 |
7,0 |
19,9 |
23,9 |
50С10Е40Б |
| Полярный |
51,5 |
5,8 |
21,3 |
14,7 |
15С20Е65Б |
| Кольский |
67,7 |
8,4 |
14,2 |
8,2 |
40С40Е20Б |
| Ловозерский |
41,0 |
26,4 |
23,0 |
9,6 |
20С40Е40В |
| Верхнепонойский |
39,4 |
2,6 |
42,2 |
15,8 |
50С20ЕЗОБ |
| Среднепонойский |
43,4 |
2,6 |
41,0 |
13,0 |
ЗОС10Е60Б |
| Лотто-Туломский |
64,3 |
4,7 |
23,0 |
8,0 |
48С36Е16Б |
| Имандро-Пиренгский |
65,6 |
18,0 |
13,2 |
3,2 |
72С15Е13Б |
| Мончегорский |
55,5 |
18,0 |
7,8 |
19,0 |
ЗОС50Е20В |
| Хибинский |
28,0 |
12,0 |
8,0 |
52,0 |
50С20ЕЗОБ |
| Верхнеумбский |
52,8 |
12,2 |
31,0 |
4,0 |
40С35Е25Б |
| Алакурттинский |
74,3 |
3,5 |
15,2 |
7,0 |
40С40Е20Б |
| Кандалакшский берег |
61,0 |
11,0 |
25,0 |
3,0 |
56С36Е8Б |
| Среднеумбский |
53,7 |
10,1 |
36,2 |
- |
55С40Е5Б |
| Стрельно-Варзугский |
37,9 |
1,2 |
58,1 |
2,8 |
21С62Е17Б |
| Ковдозерский |
65,3 |
26,4 |
8,0 |
- |
84С12Е4Б |
Таблица 3
Характеристика категорий земель и лесов по растительным районам
На беломорских островах и Кольском п-ове ели часто
имеют "столбообразную" форму, при которой ветви, опущенные
под острым углом вниз, образуют у земли своеобразный шатер, за что
их называют "елями в юбке". Сильно наклоненные и густо
переплетенные ветви значительно затрудняют птицам устройство на
них гнезд.
Годы с хорошим урожаем семян хвойных пород бывают на Кольском п-ове
довольно редко. Удовлетворительное плодоношение у сосны (Pinus sylvestris)
отмечают в среднем через 7-10 лет, У ели - через 3-7 лет. У рябины
(Sorbus aucuparia) сравнительно хорошие урожаи ягод бывают через
3-5 лет [96]. Интересно, что в городах рябина дает обильные урожаи
значительно чаще, что позволяет зимой кормиться на них свиристелям.
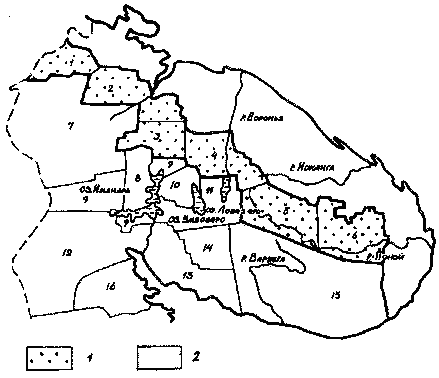
Рис. 3. Лесорастительные районы области (по В. Ф. Цветкову и В.
Г. Чертовскому, 1978):
1 - лесотундровые районы (предтундровые редколесья);
2 - северотаежные районы
Южнее полосы редкостойных лесов находятся осветленные
леса, которые в отличие от первых - воронично-лишайниковых хвойных
- называют еще воронично-зеленомошными [104]. Их северная граница
проходит восточнее оз. Пяозеро, охватывает Ковдозерскую низменность
и приозерную низменность Северной Карелии. Здесь преобладают сосняки
различных типов, занимающие 75 % лесной площади, из них зеленомошные
- около 55 %. Плотность древостоя колеблется в пределах 0,5-0,6.
В настоящее время эти леса сильно вырублены.
С продвижением от возвышенности Манселькя к Белому морю отмечается
повышение заболоченности территории. Господствующим типом болот
Северокарельской провинции является карельский аапа, который характеризуется
грядово-мочажинным комплексом в центре и сосново-кустарничково-сфагновой
растительностью по периферии [18]. Во флоре болот насчитывается
более 80 видов сосудистых растений, среди которых преобладают осоки,
молиния (Molinia caerulea) и др. На грядах господствует ерниково-молиниево-осоково-сфагновая
растительность, в мочажинах - осоково-разнотравная и разнотравно-осоково-гипновая
[103]. Благодаря широкому распространению таких болот здесь обычны
на гнездовании лебеди-кликуны (Cygnus cygnus).
Сильной заболоченностью в Карелии выделяется Прибеломорская низменность.
Три четверти ее площади занимают болота Прибеломорской провинции.
На наиболее .удаленных от моря верхних террасах находятся обширные
верховые болота грядово-озеркового комплекса, на более молодых нижних
террасах - грядово-мочажинного комплекса с изогнутыми соснами на
грядах. У самого берега моря расположены эвтрофные осоковые и тростниковые
болота [18]. По характерной растительности сфагновые грядовые болота
относятся к вереско-воронично-лишайниковым. Флора их бедна и состоит
всего из 35 видов сосудистых растений [103].
Восточнее Белого моря, начиная от Зимнего берега у устья р.Мегры,
через междуречье Северной Двины и Мезени проходит граница между
Кольско-Печорской и Валдайско-Онежской подпровинциями - редкостойными
и осветленными лесами [80]. Северная граница предтундровых редколесий
обозначена островками ели южнее б7°30' с. ш., южная - примерно вдоль
66° с. ш. [28]. Или здесь малорослые, высотой обычно 4-6 м, реже
- до 10 м. Крона их слабо развита, сомкнутость составляет всего
0,1-0,3. Повсюду сопутствующая им береза здесь еще более малоросла.
Редколесья представлены преимущественно зеленомошными, лишайниково-зеленомошными
и долгомошно-офагновыми типами растительности, нижний ярус - обычно
голубично-багульниковыми и чернично-вороничными.
Расположенные южнее редколесий северотаежные еловые леса занимают
всю прибрежную часть восточного побережья Белого моря до Онежского
п-ова включительно. В Архангельской области они располагаются на
65 % всей покрытой лесами площади. Наиболее распространены ельники-зеленомошники,
растущие на богатых дренированных почвах. Видовой состав травянистых
растений в них, так же как и подлесок, сравнительно беден. Кустарничковый
ярус, состоящий из черники, вороники, брусники, голубики, напротив,
достаточно богат [24]. Часто встречаются долгомошные и сфагновые
ельники со значительной примесью березы, которой в первую очередь
зарастают места пожарищ и вырубок [46].
Сосновые боры занимают около 25 % лесной площади Архангельской области.
Они расположены в основном по прибрежным мореким низинам с бедными
почвами. Во втором подъярусе местами произрастает ель, а на карбонатных
породах лиственница (Larix sibirica). Подлесок и травянисто-кустарничковый
ярус развиты довольно слабо. Наиболее распространенный тип сосняка
- лишайниковый с сомкнутостью крон 0,2-0,4. Большую площадь занимают
также заболоченные сосновые леса, особенно сфагновые [52].
Широко распространенные в северной тайге черника и брусника, ягоды
которых играют существенную роль в питании тетеревиных (Tetraonidae)
и других птиц, имеют высокий средний балл урожайности. В Мурманской
области он составляет 3-5 по 6-балльной шкале Каппера-Формозова,
а повторяемость таких урожаев - 8 в 10 лет. В Архангельской области
средний балл урожайности составляет 3,0-3,8, а повторяемость - 7
в 10 лет [10]. Средние весовые показатели составляют 500-1500 кг/га
черники при урожайности в 3-5 баллов и проективном покрытии 50-100
%. По В. Г. Туркову и Н. А. Шишкину [92] они составляют: черники
- 50-140 кг/га, а брусники 60-230 кг/га (средние многолетние данные
по Архангельской, Вологодской областям и Коми Республике). Естественно,
что в зависимости от характера почвы, количества света, тепла и
влаги биохимический состав ягод будет различным [39, 94]. Тетеревиные
птицы наиболее часто встречаются в тех участках леса, где урожайность
ягод брусники и черники достаточно высока и где, кроме того, обеспечена
их безопасность от хищных животных [85].
Нельзя не упомянуть также о растительности лугов, особенно пойменных.
Для тундровых пойм характерным является образование первичных лугов
из вейника (Calamagrostis langsdorffii). Из вторичной растительности
в редко заливаемых местах обычны овсяницы (Festuca rubra F. ovina)
и разнотравье, среди которого господствующее место занимает моховой
покров. На среднем уровне поймы распространены лисохвостовые Alopecurus
sp.) луга, ниже - узкие полосы канареечника ?halaroides arundinacea).
Южнее, в пойме Северной Двины, у Холмогор, на высоких уровнях сохранились
травяные еловые и березово-еловые насаждения высотой до 20-23 м,
с примесью рябины в древостое и -подлеском из жимолости (Lonicera
pallasii), черемухи (Padus avium), шиповника (Rosa pisiformis),
смородины (Ribes rubrum) и ив (Salix sp. sp.). Эколого-динамичный
ряд поймы рек северо-запада Русской равнины [58] представлен ивняком
(Salix viminalis, S. acutifolia), ельниками-пихтачами (Picea abies
x P. obovata, Abies sibirica), а также заболоченными лесами, состоящими
из ольхи (Alnus incana), березы (Betula pubescens), ивы (Salix myrsinifolia)
и ели (Picea Abies x P. obovata). Луга на высоких уровнях поймы
мелкотравно-злаковые. В травостое часто преобладают овсяница (Festuca
rubra), тимофеевка, (Phleum pratense), полевица (Agrostis gigantea).
Значительное место занимает разнотравье и бобовые - поповник (Leucanthemum
vulgare), василистник (Thalictrum minus), колокольчик (Campanula
glomerata), погремок (Rhinanthus vernalis), клевер (Trifolium pratense,
T. repens) и астрагал (Astragalus danicus). Ha средних уровнях располагаются
наиболее ценные луга, где Преобладают крупные злаки: костровые (Bromus
sp.), пырейные (Agropiron sp.), белополевицевые (Agrostis sp.),
лисохвостовые (Alopecurus sp.), а также овсяница (F. pratensis),
вейник, бобовые (Leguminosae) и крупное разнотравье. На низких уровнях
-канареечниковые и остроосоковые [58].
Высококачественные луга поймы Мезени ниже устья Пезы по урожайности
и видовому составу соответствуют заливным лугам Холмогорского района.
Многие из трав потребляются гусями во время осеннего пролета. К
сожалению, в результате интенсивного освоения поймы претерпели значительные
изменения: леса вырублены, земли распаханы.
На побережьях островов и материка Белого моря большие площади занимают
приморекие луга. В зависимости от воздействия на них морекой воды
их делят, на луга низкого, среднего и высокого уровня [57]. Растительность
примореких лугов Кандалакшского залива изучалась И. П. Бреслиной
[19].
На лугах низкого уровня, которые ежедневно в прилив покрываются
водой, она выделила 2 пояса: пояс солеросов с ассоциациями (Salicornia
pojarkovae и S. europaea), морекой астры (Tripolium vulgare) и морекого
подорожника (Plantago maritima) и расположенный на илистом грунте
с застаивающейся водой пояс бескильницы ползучей (Puccinellia phryganodes)
включающей ассоциации осоки обертковидной (Сагеx subspathacea),
ситника чернобурого (Juncus gerardii), ситняга северного (Eleocharis
septentrionalis) и блисмуса рыжего (Blysmus rufus).
На лугах среднего уровня, которые большей частью затопляются лишь
в высокие (сизигийные) приливы, различают более 8 ассоциаций. Из
них упомянем только заросли лисохвост тростниковидного (Alopecurus
arundinaceus) и колосняка песчаного (Leymus arenarius), где охотно
гнездятся утки, а также пырея ползучего (Elytrigia repens) и лебеды
(Atriplex nudicaulis A. lapponica). Семенами последней осенью постоянно
кормятся юрки (Fringilla montifringilla) и другие виды.
Луга высокого уровня испытывают воздействие морcкой воды в виде
подтопления и штормовых брызг, но происходит это редко и нерегулярно
- их заливает только во время сизигийных приливов. Растительность
лугов обычно разнотравно-злаковая. Здесь растут чина морекая (Lathyrus
aleuticus), которую охотно поедают останавливающиеся на осеннем
пролете гуменники (Anser fabalis), и канареечник тростниковидный
(Phalaroides arundinacea), служащий хорошим укрытием для птенцов
линяющих уток. В кутах губ встречаются густые заросли тростника
(Phragmites australis).
Разнообразны травостои заливных примореких лугов Онежского и Двинского
заливов Белого моря. .В их состав входя тростник (Phragmites australis),
камыш (Scirpus lacustris) полевица (Agrostis straminea), триостренник
(Triglochin maritimum), осоки (Carex glareosa, C. mackenziei) и
другие растения. На них кормятся пролетные пластинчатоклювые, некоторые
кулики, хищные, совы и воробьиные птицы.
Особо следует сказать о растительности морских островов. На Мурмане
все острова находятся в тундровой зоне и покрыты своеобразным сухим
торфом мощностью до 1 м, а иногда больше. Основной растительностью
здесь являются вороничники, местами развиваются тундрообразные травянистые
приморекие ценозы (деренник, щавелевый морошечник, папоротниковые
поросли и др.). На о-ве Большом Айнове распространены приморекие
луга из крупнотравья и колосняковые кочкарники. На скалах и литорали
растут галофиты [20].
Большинство островов Белого моря покрыты зональной северотаежной
растительностью. В зависимости от величины острова, слагаемых его
пород, удаленности от материка и других факторов И. П. Бреслина
[20] различает в Кандалакшском заливе 4 основных островных ландшафта:
- острова в губах;
- острова близ открытого или слабо изрезанного побережья материка;
- острова, удаленные от материка;
- о-в Великий, слабо отчлененный от берега узкими проливами, ландшафты
которого сходны с материковыми.
Местное население выделяет в первую очередь острова, покрытые таежной
растительностью, и луды, на которых таежная растительность отсутствует.
Ландшафт большинства островов площадью до 100-500 га довольно однообразен,
что не могло не отразиться на видовом составе птиц, который значительно
беднее, чем на материке. Мелкие и удаленные от материка острова
обычно покрыты растительностью скал, примореких лугов и вороничниками,
более крупные из них частично поросли березовым криволесьем. Вороничники,
являясь интерзональной растительностью, широко распространены и
на лудах. Разбросанные по вороничнику кусты стелющегося можжевельника
(Juniperus sibirica) и других древесно-кустарниковых пород часто
служат хорошим укрытием для птиц.
Большое скопление птиц как на маленьких, так и на более крупных
мореких островах оказывает заметное влияние на растительность, которое
наиболее сильно выражено на птичьих oбазарах Мурмана и на скалистых
лудах Белого моря, в небольших понижениях которых - в "скальных
ваннах" - Скапливается значительное количество птичьего помета
и других элементов жизнедеятельности птиц. Здесь пышно разрастается
лисохвост тростниковидный, песчаный, колосняк, лигустикум шотландский
(Ligusticum scothicum), лебеда (A. nudicaulis) и некоторые другие
виды. Нередко в таких местах держатся выводки кряквы и даже линяющие
взрослые птицы. Менее значимо влияние птиц на растительность примореких
лугов и выпуклых скальных поверхностей. Воздействие птиц на развитие
растительности на мореких островах Кольской Субарктики подробно
исследовано и описано [20].
Кольско-Беломорекий регион включает в себя юго-западную часть Баренцева
моря и все Белое. Эти моря относятся к нижнеарктической мелководной
подобласти Арктической области. С юго-запада узкой полосой вдоль
Скандинавии в нее проникают воды Бореальной области. Относительно
опресненная и теплая южная Рупиновская ветвь Нордкапского течения
переходит в Мурманскую прибрежную ветвь, которая вдоль Терского
берега заходит в Воронку Белого моря и теряется южнее, в Горле.
Акватория Баренцева моря между Восточным Мурманом и южным островом
Новой Земли находится под влиянием мористой ветви Мурманского течения
(рис. 4), от которой в свою очередь отходят Канинское, Колгуево-Печорское
и Новоземельское течения [40, 6, 7].
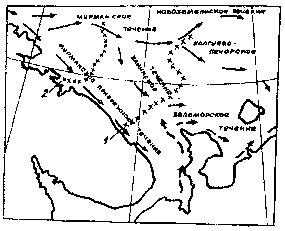
Рис. 4. Схема морских течении южной части Баренцева моря и граница
льдов в средние (1] и суровые (2) зимы
Баренцево море - относительно глубокий водоем, 77,1 % его площади
имеет глубины более 100 м. Только у Кольского п-ова вдоль берега
тянется узкая полоса небольших глубин. Кроме того, значительное
по площади мелководье - Канино-Колгуевское - расположено между Восточным
Мурманом и южным островом Новой Земли. На нем преобладают биоценозы
жестких песчаных грунтов с многочисленными двустворчатыми моллюсками.
Прибрежье Мурмана сложено главным образом гнейсами и гранитогнейсами
и находится в зоне интенсивного движения воды, вызываемого постоянным
и переменным приливно-отливным течениями и прибоем. Однако изменчивость
и скорость течений в разных местах у берега и на разной глубине
до сих пор мало исследованы. Температура воды у поверхности держится
с декабря по май в пределах от -1° до +3,5°, а к августу может повышаться
до 15-16° у Айновых о-вов (западнее п-ова Рыбачьего) и до 12-13°
у Дальнезеленецкой губы Восточного Мурмана. Соленость воды у побережья
меньше, чем вдали от него, и составляет в среднем около 33-34 %о,
достигая минимума в июне, а максимума - в декабре-апреле. Наибольшая
концентрация биогенных элементов - фосфатов и нитратов, необходимых
для развития фитопланктона и водорослей, отмечается в январе-марте,
наименьшая - в летне-осенние месяцы [40].
Юго-западная часть Баренцева моря является северо-восточным краем
высокопродуктивной зоны Северной Атлантики, протянувшейся от Ньюфаундлендской
мели к берегам Скандинавии и Новой Земли. В открытых частях моря
биомасса зоопланктона составляет в среднем 140 мг/м3, что в 3 раза
больше, чем в заливах, где его количество определено в 45 мг/м3.
В юго-западной части биомасса планктона колеблется от 200 до 2000
мг/м3, иногда достигая 8000-9000 мг/м3. Таким образом, обилие зоопланктона
здесь лишь немного уступает богатым районам Северной Атлантики [40].
Основную массу зоопланктона составляют веслоногие ракообразные (Copepoda),
главным образом Calanus finmarchicus. Этим рачком кормятся преимущественно
плактонофаги - рыбы, птицы и китообразные. В свою очередь, сельдь
(Clupea harengus), мойва (Mallotus villosus) и другие рыбы, питающиеся
планктоном, служат пищей некоторым мореким птицам, например кайрам
(Uria sp.) и моевкам (Rissa tridactyla).
Животное население дна Баренцева моря у Мурманского побережья тоже
весьма богато и насчитывает более 1,5 тыс. видов беспозвоночных.
Из них полихеты (Polychaeta), ракообразные Crustacea), моллюски
(Mollusca) и иглокожие (Echinodermata) представлены 662 видами,
что составляет 44 % от общего числа беспозвоночных [41]. Средняя
биомасса бентоса открытых частей сублиторали на глубине до 100 м
равна 311 г/м2, на глубине 100-200 м она уменьшается до 168 г/м2,
свыше 200 м - составляет менее 100 г/м2. В Мурманском прибрежье
наибольшая средняя биомасса бентоса - более 300 г - отмечена у п-ова
Рыбачьего, где преобладают губки (Porifera) и плеченогие (Brachiopoda).
От Нордкапа на Мурман распространяется биоценоз крупных моллюсков
Modiolus modiolus - Chlamys islandicus или Mactra elliptica. Вдоль
всего Восточного Мурмана биомасса бентоса составляет в среднем 75-100
г/м2. В крупных заливах при хорошей аэрации его масса значительно
больше [40].
М. В. Пропп [77] выделяет в прибрежной сублиторали 4 группы донных
сообществ: ламинариевые водоросли (Laminariales), известковые водоросли
(Lithothamnion), прикрепленные животные и сообщества мелких водорослей,
развивающихся в отдельные сезоны. Биомасса отдельных видов беспозвоночных
здесь обычно не превышает 100 г/м2.
С учетом различий в гидрологическом режиме, рельефе, поверхностных
осадках и преобладания трофических группировок бентоса выделяют
3 основных типа южных прибрежных ландшафтов Баренцева моря.
Тип А - придонные течения больше 20 м/с, грубообломочные отложения,
перенос песка и вынос алевритопелитовых фракций;
из беспозвоночных преобладают сестонофаги.
Тип Б - течения 5-20 м/с, отложения песчаные, осаждение пелитовых
фракций преобладает над переносом; наиболее распространенные животные
- собирающие детритофаги.
Тип В - течения меньше 5 м/с, отложения песчано-илистые, осаждение
алевритопелитовых фракций; из беспозвоночных преобладают грунтоеды
[89].
Литораль значительно разнообразнее по своим характеристикам, чем
сублитораль. В ее морфологии выделяют чистый илисто-песчаный пляж,
пляжи с барьером из камней в верхнем, среднем или нижнем горизонте,
а также пологую и крутую скалистую часть. Литораль Мурмана относят
к субарктической литоральной провинции [34] и выделяют 6 ее основных
биономических типов:
1. Спокойные, защищенные от прибоя бухты близ открытого океана.
2. Узкие проливы с сильным течением, также защищенные от прибоя.
3. Открытый прибойный берег.
4. Удаленные от океана глубокие куты губ без прибоя и течений.
5. Такие же губы, но сильно опресненные.
6. Эстуарии.
Следует отметить, что опреснение вызывает опускание литоральных
форм в нижние горизонты. Прибой и сильное течение, наоборот, способствуют
поднятию всех форм в более высокие горизонты. В каждом биономическом
типе обычно хорошо различимы 3 основные фации. Для фации скал характерны
заросли фукоидов с населяющей их подводной фауной, поселения мидий
(Mytilus edulis) и балянусов (Balanus sp.). На каменистых россыпях
к ним добавляется фауна боковых и нижних сторон камней, а также
ютящиеся между камнями животные. Особенностью мягких грунтов пляжей
является наличие зеленых и нитчатых бурых водорослей (Phaeophyta),
роющих червей и двустворчатых моллюсков [34]. В заветренных местах
со спокойным гидрологическим режимом собираются обитатели илисто-песчаной
литорали, средняя биомасса которых составляет 4825 г/м2, из них
4621 г (96 %) приходится на долю мидий [41]. На втором месте по
численности и биомассе стоит макома (Macoma sp.) - 115 г/м2,2 %
общей биомассы.
Обилие пищи, наличие небольших островов и обрывистых скалистых берегов
с уступами, недоступных для наземных хищников, позволяют гнездиться
на Мурмане множеству мореких птиц. Как и на Новой Земле, здесь расположены
многочисленные птичьи базары, заселенные толстоклювыми и тонкоклювыми
кайрами (Uria lomvia, U. aalge), трехпалыми чайками-моевками, Тупиками
(Fratercula arctica), а на островах Земли Франца-Иосифа и Шпицбергена
- и люриками (Plautus alle). Все эти виды питаются в основном пелагическими
рыбами, ракообразными и Другими беспозвоночными, в меньшей степени
- придонными животными. Пищей для обитателей Мурмана являются сельдь,
песчанка (Ammodytes hexapterus) и мойва, реже - треска (Cadus morhua),
у Новой Земли - сайка (Boreogadus saida) и треска [12].
Птицы, питающиеся донными беспозвоночными и рыбами, добывают их
на разных глубинах: морекие утки - до 20-50 м, чистик (Cepphus grulle)
и гагарка (Aica torda) - до 50-60 м, а не связанные с прибрежьем
тупики и кайры - больше 100 м [105, 106]. Придерживаясь бореальных
вод, на Мурман залетают птицы, характерные для Атлантического океана,
- глупыши (Fulmarus glacialis), олуши (Sula bassana) и др.
В последние годы запасы ряда видов массовых рыб в Баренцевом море
значительно сократились. Это произошло главным образом в результате
их перепромысла, а также, видимо, влияния сейсморазведки залежей
нефти и загрязнений, как образующихся на месте, так и приносимых
течениями из Атлантики. В результате произошло нарушение естественных
биоценозов и создалось катастрофическое положение в биосистеме моря.
Белое море является типичным арктическим водоемом. Его площадь составляет
90 тыс. кв. км, средняя глубина - 67 м* , .объем - 6 тыс. куб. м
[38], из которых на верхние слои приходится: 77 % на слой до 100
м и 54 % - до 50 м [91].
Периодически высказывается мнение, что Воронку с Мезенским заливом
следует отнести не к Белому морю, а к Баренцеву, так как по гидрологическому
режиму они "представляют собой не что иное, как обыкновенный
залив Баренцева моря" [9]. Так, еще Н. Данилевский [36] писал,
что поморы считали границей Белого моря мыс Орлов-Терский, севернее
которого начинается, по их мнению, "океан", т. е. Баренцево
море. "Там и волна становится океанская, и фауна не беломорская,
даже нередко заплывают киты". Действительно, Мурманское прибрежное
течение от поверхности до дна сплошным потоком шириной в 25-35 км
проходит вдоль Терского берега до мыса Городецкого. Южнее его значительная
часть вод поворачивает обратно и уходит на север (рис. 5). Таким
образом, западная часть Воронки Белого моря оказывается заполненной
баренцевоморской водой [91].
В свою очередь, поступающая из Баренцева моря часть потока в Горле
Белого моря интенсивно перемешивается турбулентным течением и теряет
свои свойства. Так гидрологический режим Горла, являясь непроходимым
экологическим барьером .[32], препятствует проникновению в Бассейн
Белого моря баренцевомореких форм [91]. И все же, несмотря на это,
мы придерживаемся более распространенного взгляда, что северная
граница Белого моря проходит между мысами Святой Нос и Канин Нос.
Ежегодно (преимущественно зимой) через Горло ''Белого моря в его
Бассейн вливается около 2000 куб. км баренцевоморекой воды, которая
заполняет глубоководный желоб, протянувшийся на северо-запад к Турьему
мысу в Кандалакшском заливе. Много воды поступает в Белое море и
из рек -
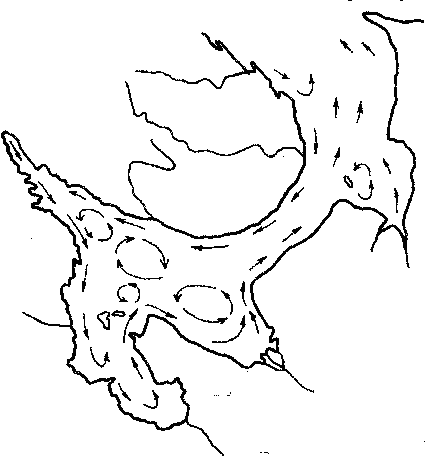
Рис. 5. Схема постоянных течений Белого моря
около 237 куб. м. Этот пресный сток образует постоянное течение,
которое под влиянием силы Кориолиса направляется (против часовой
стрелки) вдоль берегов Бассейна и его заливов и создает у восточных
берегов сточное течение в Баренцево море. Его годовой объем - около
2200 куб. м. Зимой выносу льда через Горло Белого моря способствует
также муссонный юго-западный ветер.
Гидрологи считают, что водообмен Белого моря с Баренцевым имеет
для первого большое значение, так как способствует ежегодно (или
один раз в два года) происходящему в нем процессу обновления вод
[38, 42, 91], а также насыщению кислородом глубинных вод, которые
каждый год обновляются на 2/3[37].
Кроме кругового постоянного течения у устьев заливов в Бассейне
Белого моря отмечаются циклонические (против часовой стрелки) круговороты
воды (см. рис. 5). Самый большой из них, в котором, кстати, расположен
"полюс холода" моря, находится у устья Двинского залива,
поменьше - у Кандалакшского залива и самый маленький - северо-восточнее
Соловецких о-вов. Между этими круговоротами происходит антициклоническое
вращение воды с "полюсом тепла" [91]. Круговороты у устьев
заливов, как считает 3. Г. Паленичко [73], создают определенную
преграду для распространения пелагических личинок, что способствует
развитию местных адаптивных признаков у обитающих там форм.
Как и во многих других морях, покрывающихся на зиму льдом, в Белом
море находятся стационарные полыньи, которые поддерживаются у западных
берегов отжимными ветрами (рис. 6). Вследствие конвекции вод зимой
и более раннего начала их прогрева весной в весенне-летний период
на месте полыней может оставаться пятно относительно теплых и богатых
биогенами вод на поверхности и более холодных и соленых у дна. Все
это способствует повышению биомассы фито- и зообентоса, скоплению
рыб, млекопитающих и птиц. Повышенную биомассу и раннее (одновременное
с Западным Мурманом) развитие планктона гидробиологи отмечали у
северо-западной оконечности Терской полыньи в районе Святого Носа.
И это несмотря на то, что температура воды здесь ниже почти на 3°.
Наличие в Белом море стационарных полыней позволяет не только зимовать
ряду перелетных водоплавающих птиц, но и существовать оседлым беломореким
популяциям обыкновенной гаги и атлантического чистика [56].
Рис. 6. Схема деления Белого моря:
/ - стационарные полыньи (по В. Н. Купецкому. 1961)
Несмотря на незначительные размеры, Белое море настолько разнообразно,
что гидробиологи отмечают особые биономические типы Воронки, Мезенского
залива. Горла, Бассейна, Кандалакшского, Онежского и Двинского заливов
(см. рис. 6). Этому способствуют различия в глубине и рельефе дна,
очертаниях берегов, температуре и солености воды, течениях и ряд
других характеристик. В результате не только биоценозы разных частей
моря, но и некоторые населяющие их виды растений и животных значительно
отличаются по ряду признаков [33].
Как среду обитания птиц Белое море нет необходимости делить так
дробно, как это делают гидробиологи и другие специалисты. Достаточно
разделить его на 3 части: западную (шхерную) с Кандалакшским и Онежским
заливами, восточную (открытую), куда входит большая часть Бассейна
и Двинский залив, и северную (внешнюю), которая состоит из Мезенского
залива, Горла и Воронки Белого моря.
Настоящих морских птиц - представителей отрядов трубконосых (Procellariiformes),
веслоногих (Pelecaniformes) и других, использующих в пищу преимущественно
пелагических животных, - в Белом море мало. В незначительном количестве
они залетают в Воронку Белого моря, изредка проникая в его внутренние
части. Основная масса беломореких птиц относится к прибрежным. Они
питаются главным образом донными животными, обилие которых здесь
зачастую больше, чем в других северных морях [69]. Недаром Белое
море иногда называют морем моллюсков [73]. Донные животные доступны
для многих птиц на литорали и в верхней сублиторали до глубин 5-15
м, где, кстати, и находятся наиболее богатые биоценозы бентоса.
Особенно надо выделить шхерные районы с их мозаичным расположением
и разнообразием биоценозов. Безопасное гнездование прибрежным птицам
обеспечивают многочисленные маленькие островки, на которых отсутствуют
наземные хищники.
Учитывая, что сублитораль небольших губ является основным местом
кормежки птиц, отметим, что размещение различных беспозвоночных
мягких грунтов зависит, кроме других особенностей, и от степени
крутизны дна, а эпифауны - от наличия твердого субстрата и хорошей
гидродинамики [72]. Что же касается запасов корма на беломорекой
литорали, то здесь гидробиологи выделяют 9 биотопов: скалистый берег;
валунно-глыбовая литораль; песчаная литораль; илистая литораль;
прикрепленные фукоиды; пелагиаль над литоралью; пена, плавающие
фукоиды, пленка воды; штормовые выбросы, супралитораль; лед и снег
[50].
Воронка Белого моря испытывает прямое влияние Баренцева моря. Приливная
волна попадает через нее в Мезенский залив, где, увеличиваясь на
мелководье, создает у берегов значительную разницу в уровне воды.
Так, во время прилива и отлива она составляла в среднем 3,1-4,6
м, в устье р. Мезени - 6,6 м, а в сизигии - больше 10 м [60].
Берега Воронки, Горла и Мезенского залива относительно высокие.
Терский берег имеет узкую литораль, скалист и постепенно понижается
к югу. Он изрезан небольшими губами, среди которых выделяется неглубокий
Лумбовский залив с неровным дном, прикрытый со стороны моря островами
и, как пишет 3. Г. Паленичко [73], "исключительно богат крупными
бурыми водорослями", где "вырастает 10-метровая ламинария".
Канинский берег в северной части обрывист и скалист. Южнее, от Торны
до р. Кии, он образует полосу дюн, достигающую вблизи рек ширины
в несколько сотен метров. Дальше к югу высота берега неоднократно
меняется, то повышаясь, то понижаясь.
Средняя глубина Мезенского залива составляет 13 м. Рельеф дна постоянно
меняется под действием штормов и весьма значительных приливно-отливных
течений. Сточное беломорекое течение, идущее вдоль восточного берега,
усиливается севернее устья водами Мезени. Е. Ф. Гурьянова [33] делит
акваторию залива на .2 района - эстуарный, широкой полосой протянувшийся
от мыса Нерпинского у устья Кулоя на север вдоль Конушинского берега,
и морекой, расположенный на западе (рис. 7). Литораль эстуарного
района очень широкая, местами доходящая до 10 км. Ее образуют чистые
пески, на поверхности которых можно встретить тонкий слой размельченного
торфа, выносимого почвенными водами. В устьях многочисленных рек
песок покрыт толстым слоем няши - полужидкого глинистого ила, который
смывается сильными штормами, чтобы снова 'oнакопиться в штиль. Выносимая
реками минеральная взвесь резко понижает прозрачность воды. Заселяющая
районы Мезенского залива фауна неоднородна: в одном месте она солоноватоводная
эстуарная, в другом - морекая. Зарослей водорослей и полей зостеры
(Zostera sp.) в Мезенском заливе нет, планктон очень беден. В его
составе отмечено [51] Только 29 видов биомассой, не превышающей
40,4 мг/м3. Бентос также незначителен, представлен 165 видами, в
основном
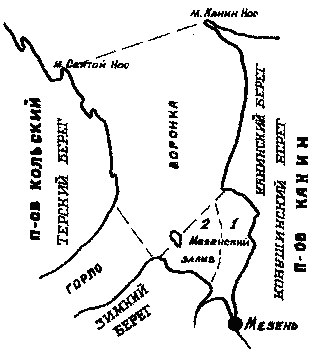
Рис. 7. Деление Мезенского залива на эстуарный (1) и морской (2)
районы
беспозвоночньми - фильтраторами (Modiolus, Tridonta, Mytilus) и
собирающими детритофагами (Macoma). Большая часть Мезенской губы
занята сообществами сидячих фильтратов, в частности биоценозом Modiolus
modiolus + Balanus crenatus общей биомассой 363,2±183,7 г/м2 (М.
modiolus - 180.7±155.2 г/м2 и В crenatus - 105,0±88,9 г/м2). Близ
устьев Мезени и Кулоя в сублиторали располагаются биоценозы Tridonta
borealis общей биомассой 35 г/м2 и Macoma balthica . 182,0 г/м2
при биомассе ведущего вида - 149.0 г/м2 [5, 66]. На нижнем горизонте
литорали разбросаны пятна Arenicola manna, а ниже их -обширные поселения
Macoma balthica. Наиболее плотные поселения макомы отмечены в районе
мыса Стрельничного Абрамовского берега, где моллюски достигают особо
крупных размеров [33]. В устье Мезени литораль покрыта толстым слоем
безжизненной няши. В прилив сюда поднимаются с водой крабы Hyas
araneus) и креветки (Crangon crangon). В целом общая биомасса бентоса
Мезенского залива составляет в среднем около 300 г/м2, что почти
вдвое превышает биомассу в Горле Белого моря [66].
Терский, Зимний и Летний берега Бассейна и Двинского залива имеют
пологие очертания, около них почти нет островов. Относительное однообразие
здесь нарушают только дельта Северной Двины и Унская губа. Выносимое
Северной Двиной огромное количество пресной воды распресняет море,
постоянным течением направляет вдоль Зимнего берега к Горлу Белого
моря. Малакофауна Двинского залива довольно бедная как в количественном,
так и в качественном отношении. Общая биомасса ее в среднем на порядок
меньше, чем на таких же глубинах в Кандалакшском и Онежском заливах
[95]. Распределение бентоса довольно однотипно. Песчаная литораль
лишена макроформ. Верхняя сублитораль до глубины 5 м заселена сообществом
единственного массового вида - мидии, средняя биомасса которого
колеблется от 1 до 10 кг/м2. Ниже находится полоса с мизерным населением
общей биомассой менее 10 г/м2. Глубже 10 м располагаются биоценозы
полихет и двустворчатых моллюсков биомассой до 150 г/м2.
У Летнего берега от Ухтнаволока до д. Лопшеньги сообщество мидий
постепенно сменяется биоценозом ламинарии биомассой 10-20 кг/м2.
Пролетные нырковые утки останавливаются здесь осенью на непродолжительный
отдых и кормежку. В северо-восточной части залива растет много макрофитов
и встречаются поселения Arenicola marina, которые спускаются до
глубины 5 м [67].
Унская губа, соединенная с Двинским заливом узким проливом, довольно
обширная по площади, но мелководная - до 15 м глубиной. В ее сублиторали
обычно можно встретить мидиевые поселения биомассой до 3 кг/м2,
а на литорали располагаются сообщества Hydrobia ulvae общей биомассой
75 г/м2 [67].
Глубоководный Бассейн Белого моря значительно меньше по сравнению
с другими его частями используется птицами. Узкая на значительном
протяжении Терского берега литораль заметно расширяется близ устья
р. Варзуги и восточнее р. Чаваньги. В макробентосе каменистой литорали
восточнее устья р. Варзуги доминируют Fucus vesiculosus и Mytilus
edulis общей биомассой около 5 кг/м2, а в верхней сублиторали -
биоценоз губок Halichondria panicea биомассой около 200 г/м2 [68].
В прибрежной полосе восточнее р. Чаваньги постоянно обитают (линяют,
зимуют и останавливаются на пролете) нырковые утки [14]. Материковое
мелководье здесь относительно широкое, на нем разбросано много подводных
камней, банок и корг, омываемых быстрым течением; грунт каменист.
В Бассейне наиболее часто можно встретить залетающих во внутренние
части моря представителей отряда трубконосых.
Западная шхерная часть Белого моря больше, чем другие .районы, заселена
птицами. Обширные мелководья Онежского залива и Карельского побережья
сочетаются здесь со значительными глубинами центрального желоба
и перепадами глубин Кандалакшского залива. Более двух тысяч небольших
островов, часть которых сгруппирована в Кандалакшские, Кемские,
Сумские и Онежские шхеры, тесно связаны с причудливо изрезанными
Карельским и Поморским берегами, образуя сложное кружево с большим
разнообразием экологических условий. Многочисленные реки выносят
большое количество пресной воды, которая оказывает заметное влияние
на формирование верхнего, наиболее изменчивого и насыщенного жизнью
слоя воды.
Онежский залив более мелкий, чем Двинский и Кандалакшский. В его
открытой части глубины изменяются преимущественно от 20 до 50 м.
Вершина залива южнее д. Пурнемы и о-ва Ковдострова почти вся лежит
в пределах 20-метровой изобаты. Обширная акватория Онежских шхер
и прибрежная 5-километровая полоса имеют глубины менее 5 м. Приливно-отливные
течения достигают в сизигии скорости до 1 м/с и более. Наибольшая
разница в уровне воды между приливом и отливом составляет: у Соловецкого
архипелага - около 1 м, а в куту Онежского залива - около 3 м. Постоянное
течение направляется в залив с севера через Западную Соловецкую
салму, затем вдоль Поморекого берега до кута и как сточное течение,
усиленное водами р. Онеги, выходит в Бассейн Белого моря через Восточную
Соловецкую салму. В середине залива образуются круговороты различной
направленности. Общая картина течений осложняется не только рельефом
и речным стоком, но и глубинными компенсирующими противотечениями.
Вследствие небольших глубин и частых штормов вся масса воды в Онежском
заливе перемешивается и имеет одинаковую температуру и соленость.
Средняя температура воды у берега составляет в июле-августе 15-18°,
у Соловецкого архипелага - 10°. Соленость колеблется в среднем от
20 %о в апреле до 29 %о в январе [8]. Приустьевые участки рек заметно
опреснены. Следует напомнить, что степень опреснения зависит от
интенсивности пресного стока, рельефа дна, конфигурации берега,
мореких течений и ветрового волнения. При сравнении в июне-июле
1952 г. характеристик вод приустьевых участков моря у рек Выга с
Шижней и р. Нюхчи наиболее значительное различие было отмечено в
солености воды, а по другим физико-химическим показателям они были
сходны [59].
К Поморекому берегу Онежского залива приурочено множество островов,
большая часть которых входит в Кемские, Сумские и Онежские шхеры
(рис. 8). Каждый из них имеет свои характерные черты. В шхерах и
у сильно изрезанного берега как на литорали, так и в верхней сублиторали
преобладают разнообразные мозаично расположенные биоценозы. Здесь
выделяют 3 типа подводных ландшафтов [90].
Тип А. Распространен шире других. Донные отложения состоят из гравийно-галечного
материала (глубже 6-7 м - пески). Среди водорослей доминируют Ahnfeltia
plicata, Fucus vesiculosus, F. serratus.
Тип Б. Располагается вдоль берега узкой полосой в 30-40 м, занимая
обширные площади с грядами. Это грубообломочный валунно-гравийно-галечный
субстрат, поросший F. vesiculosus и F. serratus.
Тип В. Характеризуется слабой гидродинамикой. Грунты песчано-глинистые
и глинистые, водорослей мало, только кое-где растут фукусы.
Вследствие того, что вода в Белом море не столь прозрачна, как в
Баренцевом море, водоросли растут здесь преимущественно до глубины
12-13 м, а в губах с еще меньшей прозрачностью - до в м. Это широко
распространенные Ascophyllum nodosum, фукусы (Fucus vesiculosus,
F. serratus), ламинарии (Laminaria saccharina, L. digitata), анфельция
(Ahnfeltia plicata) и многие другие. Наибольшее разнообразие водорослей
отмечается в Соловецком архипелаге - 158 видов, у Терского, а также
в южной части Карельского и северной части Поморекого берегов -
по 142-143 вида, в то время как в Мезенском заливе и у Зимнего берега
встречено всего по 86-89 видов.
Рис. 8. Схема расположения островов в Онежском заливе
У западных берегов Белого моря биомасса водорослей в 2-3 раза больше,
чем у восточных [21]. Наличие крупных водорослей усложняет подводный
ландшафт, а также способствует увеличению видового состава обитателей
подводного мира. Обычно над грунтом с обломками скал, камней и заиленным
песком колеблются пластины ламинарий, которые используются различными
беспозвоночными как твердый субстрат. Таким образом, здесь могут
обитать животные различных экологических Групп. Глубже 12-15 м водорослей
становится мало, количество ила быстро нарастает и соответственно
увеличиваются поселения характерных для мягких грунтов беспозвоночных.
На значительном протяжении вдоль Поморского берега Онежского залива
нет четкой границы берега моря. Обширная, часто в несколько километров,
песчаная или илистая литораль переходит в заболоченный приморекий
луг до 1 км шириной и более, который при нагонных и сгонных водах
то затопляется .полностью, то не заливается даже до верхнего этажа
среднего горизонта литорали. Нерегулярная затопляемость верхнего
и частично среднего горизонтов литорали в сочетании с ее значительной
шириной создают неприемлемые условия для обитания здесь многих литоральных
беспозвоночных. В заветренных тихих губах материка у Сумских шхер
накапливаются мелкие неорганические и органические частицы, образуя
няшу - насыщенную водой вязкую и липкую массу. Наоборот, у открытых
берегов течения и шторма уплотняют песок, он становится плотным,
почти как асфальт; прибрежных птиц тут гнездится мало, зато в отдельных
местах собираются тысячи мигрантов.
Обычная для шхер литораль имеет ширину в несколько десятков метров,
песчаный и илисто-песчаный грунт, по которому разбросаны различного
размера валуны. Если они достаточно большие и высокие, то лед, покрывая
их зимой, не соприкасается с грунтом. Образуется много мест, где
сохраняются различные животные и растения.
На твердых грунтах литорали обычно располагаются биоценозы с преобладанием
фукоидов, мидий, балянусов и литторин (Littorina sp.) средней биомассой
около 5500±4500 г/м2 и даже 12000±600 г/м2. На мягких грунтах представлены
.биоценозы с доминированием инфауны - Macoma balthica, Mya Tenaria,
Arenicola marina и морекой травы Zostera marina. Последняя у границы
с сублиторалью во многих местах образует сплошной пояс. Биомасса
растений и животных на мягких Грунтах колеблется от нескольких десятков
до 800-3000 г/м2. Заросли зостеры непостоянны вследствие случающегося
иногда почти полного ее отмирания, после чего восстановление идет
в точение нескольких десятилетий. В периоды расцвета, что, например,
отмечалось в середине нашего столетия, вес зостеры может достигать
5-6 кг/м2. Здесь же в массе можно встретить Hydrobia ulvae. Таким
образом, определяющим на литорали являются форма рельефа, скорости
течения и характер осадконакопления, в связи с чем выделяют ландшафты
с преобладанием абразионных и аккумулятивных форм и соответствующими
им экосистемами жестких и мягких грунтов [9].
Зимой южная часть Онежского залива и район Сумских шхер покрываются
льдом берегового припая. На большей части открытой акватории держится
дрейфующий лед, у границы с которым имеются стационарные полыньи.
Довольно многочисленны они на Сорокском мелководье между Помореким
берегом и о-вами Жужмуями. Полыньи располагаются здесь над глубинами
до 10-15 м, что позволяет успешно зимовать в них большому количеству
гаг беломорекой популяции. Встречаются полыньи также на Муксалмском
рифе у Соловецкого архипелага, в Кемских шхерах и в других местах.
Кандалакшский залив считается наиболее разнообразным по экологическим
условиям среди заливов Белого моря. С юго-востока в него входит
глубоководный желоб, в котором напротив Турьего мыса находится самая
большая впадина моря глубиной 340 м [37]. В средней части залив
мельчает, 200-метровая изобата проходит между Пильским и Кузокоцким
мысами, а 100-метровая узким языком вдается на северо-запад чуть
дальше Порьей губы. В вершине залива на площади 300 кв. км расположены
Кандалакшские шхеры (рис. 9). Вдоль изрезанного губами Карельского
берега находится множество островов различной величины.
Колебания уровня воды в приливы и отливы достигают в Кандалакшском
заливе в среднем около 2 м. Постоянное сточное течение направлено
вдоль Карельского берега. В расширенной части залива образуются
круговороты, действующие по ходу и против часовой стрелки. Поверхностный
слой воды у устья губы Чупа прогревается летом до 13-14°, а глубже
50 м - постоянно остается около 0°. Соленость в течение года колеблется
от 15 до 26 %о у поверхности и постоянно держится около 28 %о глубже
50 м [9]. Зимой значительная часть залива покрывается льдом берегового
припая, кромка которого обычно проходит в районе Турьего мыса, а
в теплые зимы - северо-западнее его. Вдоль берегового припая сохраняются
полыньи, которые тянутся вдоль Карельского берега к югу. Сложная
и постоянно меняющаяся в шхерах система струй приливно-отливных
течений подмывает лед a узких местах, а в теплые зимы поддерживает
незамерзающие .полыньи. Некоторые из них, наиболее постоянные, например
у о-ва Великого, гаги используют для зимовки. Полыньи -поддерживаются
и сильными струями отводящих каналов, гидростанций.
Рис. 9. Схема деления Кандалакшского залива:
А-Б - граница между кутом и средней частью;
В-Г - граница между средней и открытой частями
В Кандалакшском заливе отмечена наибольшая в Белом море 'биомасса
зоопланктона, которая составляет в верхнем 50-метровом слое в среднем
290 мг/м3, в то время как в Бассейне - 257 мг/м3, в Двинском заливе
- 219 мг/м3, в Онежском - 102 мг/м3, Мезенском - 11 мг/м3 и Воронке
- 16 мг/м3. Крупные пятна планктона распределяются по акватории
в зависимости от циркуляции вод. Участки в десятки квадратных километров
могут иметь повышенную концентрацию животных биомассой более 500
мг/м3 [78]. Наблюдения за зоопланктоном в губе Чупа в 1957 г. показали,
что обычными и более обильными были 4 вида: Pseudocalanus elongatus
- 268 мг/м3, Temora longicornis -201 мг/м3, Oithona similis - 178
мг/м3 и Acartia longiremis -151 мг/м3 [44].
Сложная система рельефа дна, обнажающихся в отлив мелей, островов
и затейливых очертаний берегов определяет изменчивость водных струй
приливно-отливных течений. Мозаичность экологических условий на
мелководьях сублиторали способствует развитию разнообразных биоценозов.
Собственно повсеместно в массе встречается один вид, который служит
пищей почти для всех птиц, питающихся морекими беспозвоночными,
- это мидия. Плотность поселений моллюска достигает в губе Чупа
13,5 тыс. экз./м2, а биомасса доходит до 18,6 кг/м2 [27]. Максимальная
биомасса мидий в Кандалакшском заливе достигает 50 кг/м2 [68].
По месту нахождения мидиевых поселений их делят на 3 основных типа:
1. Литоральные банки в губах и на открытых побережьях биомассой
не более 5-6 кг/м2.
2. Банки в узкостях на литорали и в сублиторали биомассой до 15-20
кг/м2.
3. Эстуарные, как правило, сублиторальные банки биомассой 20-32
кг/м2.
К перечисленным трем типам [61] следует добавить четвертый - на
голомянных, т. е. удаленных от материка, мелководьях, например в
районе Роговой луды. Разрозненные поселения мидий в виде небольших
щеток встречаются очень широко.
На заиленных грунтах в губе Чупа наибольшую биомассу имеет Муа arenaria
- 720 г/м2. Однако этот моллюск труднодостижим для птиц. Более доступными,
например, для уток являются моллюски биомассой от 130 до 500 г/м2.
Это -Macoma balthica, Modiolus modiolus, Serripes groenlandicus,
Arctica islandica, Tridonta borealis, Buccinum undatum и Musculus
fliscors [63]. Преимущественно этими видами кормятся турпаны Мelanitta
fusca). Птицы используют в пищу и различные виды р. Littorina, численность
которых даже при небольшой биомассе (меньше 100 г/м2) может достигать
сотен и даже тысяч особей на 1 кв. м. Как объекты питания для птиц
представляют интерес также полихеты pp. Nereis, Nephtys и Aphroditidae,
ракообразные pp. Gammarus, Caprella и Hyas araneus, морекие звезды
Asterias rubens.
В лагунах, т. е. прибрежных мелководных водоемах, водообмен которых
с акваторией моря затруднен, складываются своеобразные экологические
условия. Обычно с глубиной соленость воды увеличивается, а температура
понижается. На дне нередки отложения ила. Подобные условия наблюдаются
и в кутах губ, у закрытых островами участков берега со спокойной
гидродинамикой. Видовой состав бентоса здесь довольно беден, но
плотность обитания и биомасса , отдельных видов часто значительны.
Преобладающие над другими гидробионтами черви и моллюски [102] служат
пищей уткам и куликам р. Tringa, которые предпочитают питаться инфауной.
Анализируя факторы, определяющие биологическую структуру Белого
моря, К. В. Беклемишев с соавторами [11] разделил Кандалакшский
залив на открытую и кутовую части с границей между мысом Пентельским
и Кузокоцкой губой, где находится крутой свал в глубоководный желоб
(см. рис. 9). На глубинах более 100 м в открытой, внешней части
Кандалакшского залива на дне преобладают беспозвоночные собиратели
-детритофаги одного биоценоза Portlandia arctica. Биомасса бентоса
здесь мала и не превышает 100 г/м2. На меньших глубинах северо-западное
внешней части и у ее берегов экологические условия гораздо разнообразнее,
часто преобладают сестонофаги различных биоценозов биомассой до
300 г/м2, а местами в 10 и даже в 100 раз больше (рис. 10).
Кутовую часть Кандалакшского залива рационально дополнительно разделить
на два участка - на собственно кут с Кандалакшскими шхерами и на
среднюю часть залива юго-восточнее линии, соединяющей Колвицкую
и Княжую губы [11]. В" средней части большие площади заняты
биоценозами Joldia hyperborea, Balanus balanus, Macoma calcarea
и другими, а oнаибольшая биомасса бентоса приурочена к широкой полосе
акватории вдоль Карельского берега. Птицы здесь держатся преимущественно
на мелководье близ Роговой луды и Ковдской губы, а также на прибрежных
акваториях материка. В Кандалакшских шхерах тесное переплетение
различных экологических условий вызывает мелкопятнистое размещение
всевозможных групп донных беспозвоночных, а на мелководьях -практически
повсеместное наличие мидий. Значительная биомасса зоопланктона [75]
и множество маленьких островов создают благоприятные условия для
гнездования морских птиц.
Рис 10 Распределение общей биомассы бентоса в Кандалакшском заливе
(по . К В. Беклемишеву. Н. Л. Семеновой и О. И. Малютину. 1980):
I . биомасса меньше 10 г/м'; 2 . 10-30 г/м';
3 - 30-100 г/м2; 4 - 100.300 г/м";
5 - больше 300 г/м'
До 1960 г в Белом море было отмечено 56 форм (видов,
подвидов и natio) рыбообразных и рыб, не считая пресноводных, которые
заходят в сильно опресненные участки прибрежных вод [З]. Кроме того,
в октябре-ноябре 1964 г. в Кандалакшском заливе были добыты молодые
гигантские акулы (Cetorhinus maximiis), которых раньше тут не встречали.
Таким образом, теперь в Белом море известно 57 форм. Из них постоянными
обитателями являются 42 формы, остальные заходят сюда из Баренцева
моря для икрометания или откорма. Из постоянных обитателей о видов
являются эндемиками: беломорекая сельдь -Clupea harengus pallasi
natio maris-aibi, беломорекая треска -Gadus niorhua maris-aibi,
беломорекая корюшка - Osmerus eperlanus dentex iiatio dvinensis,
беломорекий ликод - Lycocles pallidus maris-aibi, беломорекая речная
камбала - Pleuronectes flesus bogdauovi [65]. Наибольшее видовое
разнообразие рыб отмечено в Воронке Белого моря, Кандалакшском заливе,
у Терского и Карельского берегов, где отмечается большая соленость
воды. В Мезенском, Двинском и Онежском заливах количество видов
меньше, но запасы некоторых из них довольно значительны, например,
сельди, наваги (Eleginuis navaga), корюшки.
Большинство видов рыб можно разделить по сроку нереста на весенне-
и зимненерестующие. Теплолюбивые весенненерестующие виды - сельдь,
корюшка, пинагор (Cyclopterus liimpus), трехиглая колюшка (Gasterosteus
aculeatus) - имеют прикрепленную икру, так как размножаются на прибрежных
мелководьях в период максимальной скорости сточного течения. Пелагическую
икру имеют только то рыбы, нерест которых происходит вдали от берега
или в период летней межени. Это -ершоватка (Limanda limaiida) и
речная камбала (Pleuronectes flesus bogdanovi). Холодолюбивые зимнонерестующие
рыбы - навага, треска, бычки (Cottidae) и другие - нерестятся в
водах баренцевоморекого течения вдали от берега. Их икра не прикрепленная,
а свободнодрейфующая. В зависимости от глубины нахождения она может
выноситься в Баренцево море поверхностными водами, как у сайки,
или задерживаться у дна, как у наваги [З]. Сравнивая видовой состав
рыб в губах Карельского и Поморекого побережий, Ф. Б. Мухомедияров
[64] отмечает, что у Карельского берега наблюдается повышенная численность
лишь тех видов, которые проявляют эврибионтность только в период
размножения весной, - трехиглая колюшка, бычки. Виды, приспособившиеся
к резким сезонным колебаниям всех основных факторов - температуры,
солености, скорости течения и др., больше распространены в Онежском
заливе (навага, корюшка).
Многолетний опыт рыболовства в Белом море показал, что запасы ряда
видов подвержены значительным колебаниям [55]. Это относится в первую
очередь к сельди, трехиглой колюшке и песчанке (Ammodytes hexapterus).
Например, несметные полчища колюшек подходили раньше к берегам на
икрометание. Они появлялись около 1 июня и держались на мелководье
почти 3 месяца. В этот период ими питались многие птицы, а в Кандалакшском
заливе колюшки являлись основным массовым кормом для полярных крачек
(Sterna paradisaea) во время их гнездования. К середине 1960-х гг.
трехиглая колюшка почти перестала появляться у берегов, и количество
крачек здесь значительно уменьшилось. В 1970-х гг. с потеплением
подходы колюшки стали восстанавливаться, но затем снова упали. Более
мелкая девятииглая колюшка (Pungitius pungitius) обычна в пресных
и солоноватых водоемах, она не бывает столь многочисленной, как
трехиглая.
Песчанка обитает только на песчаных грунтах, иногда большими косяками
появляется у поверхности воды, где выступает конкурентом сельди.
Ею главным образом кормятся гагарки. При отсутствии песчанки они
добывают мелкую сельдь, которой питаются также утки, крохали, чайки
и другие птицы.
На Сорокском мелководье осенью навагой и корюшкой кормятся многочисленные
пролетные гагары (Gavia arctica и G. stellata).
Следует отметить, что одиночных видов донных рыб в Белом море значительно
больше, чем стайных. Из них, кроме бычков, камбал (Pleuronectidae)
и люмпеновых (Lumpenidae), надо упомянуть еще маслюка (Pholis giinnellus),
которого можно найти во время отлива под водорослями на нижнем горизонте
литорали. Этими видами кормятся крохали и атлантические чистики.
Если промысловые рыбы достаточно хорошо изучены, то этого нельзя
сказать о многих непромысловых видах [3, 64, 74]. Это и понятно,
если учесть, что все 30 форм встречаются относительно редко.
Из млекопитающих в Белом море распространены белуха (Delphinapterus
leucas), кольчатая нерпа (Pusa hispida), гренландский тюлень (Pagophilus
groenlandica), морекой заяц (Erignathus barbatus). Еще совсем недавно
можно было встретить и морекую свинью (Phocoena phocoena). Из перечисленных
видов для птиц наибольшее значение, пожалуй, имеют зимние залежи
гренландского тюленя в Горле и Бассейне Белого моря, около которых
собираются некоторые виды зимующих птиц.
Сделанное описание конспективно и, безусловно, не могло вместить
сведения о всех интересных и принципиальных исследованиях, проведенных
в крае.
В заключение хочется отметить, что описанный регион является наиболее
хозяйственно освоенной северной частью нашей страны. За последние
десятилетия полунатуральный тип хозяйства сменился здесь многоотраслевой
современной экономикой, включающей добывающую и перерабатывающую
промышленность, лесное, рыболовное и сельское хозяйства. Бурные
темпы хозяйственного освоения региона не могли не сказаться на птицах.
Из гнездящихся видов сократилась численность крупных хищников, оттеснены
в малопосещаемые людьми места лебеди и гуси, гибнет значительное
количество обыкновенных гаг. Принятые для охраны птиц меры до недавнего
времени помогали поддерживать в заповеднике численность обыкновенных
гаг и птиц-базарников. Ряд видов приспособился к происходящим изменениям.
В городах гнездятся сизые голуби и воробьи, кормятся зимой большие
синицы, снегири, свиристели, а летом чайки. Обилие птиц привлекает
в населенные пункты хищников, особенно поздней осенью и зимой. Однако
разнообразие видов катастрофически уменьшается. Идет деградация
природных экосистем.
Наверх
Литература
Агроклиматическнй справочиик по Мурманской области. - Л.: Гидрометиздат,
1961. 88 с.
Алисов Б. П. Климаты СССР. - М.: МГУ, 1956.
Алтухов К. А., Михайловская А. А., Мухомедияров Ф. Б. в др. Рыбы
Белого моря Петрозаводск, 1958, 163 с.
Андрияшев А. П. Рыбы северных морей СССР. - Л.-М.: Изд. АН СССР,
1954, 566 с.
Арроиет Н. М., Бергер В. Я., Луканин В. В. и др. Донные биоценозы
Мезенского залива Белого моря//Пробл. изуч. рацион, использ. и охраны
природных ресурсов Белого моря//Тез. докл. регион, конф. - Архангельск,
1985. С. 80-81.
Арсеньева Н. Я. К вопросу о влиянии приливно-отливных течении IA
климатические и гидрологические условия моря//Тр. ГОИН, вьп;. 64.
Вопросы гидрологии северных морей. - Л.: ГИМИЗ, 1961. С. 93-102.
Атлас Мурманской области. - М., 1971. 33 с.
Бабков А. II. Гидрологическая характеристика Онежского залива Белого
моря//Исслед. фауны морей. 33 (41) Экосистемы Онежского залива Белого
моря. -Л.: ЗИН АН СССР, 1985. С. 3-10.
Бабков А. 11., Голиков А. 11. Гидрокомплексы Белого моря. - Л.:
3EH АН СССР, 1984. 104 с.
Барыкина В. В. География плодоношения дикорастущих ягодных растений
на территории европейской части СССР//Автореф. дис. канд. биол.
наук. - М., 1966. 20 с.
Беклемишев К. В., Семенова Н. Л., Малютин О. И. Факторы, определяющие
биологическую структуру Белого моря/./Биология моря. - Владивосток:
Наука, 1980, № 1. С. 8.20.
Белопольский Л. О. Состав корма мореких птиц Баренцева моря//3оол.
исслед. сев.-зап. районов СССР// Уч. зап. Калининград, гос. ун-та
Вып. VI. -Калининград, 1971. С. 41-67.
Бианки В. В. Численность и размещение массовых мореких птиц в западной
части Белого моря/./Пробл. использ. промысл, ресурсов Белого моря
и внутр. водоемов Карелии. - М. -Л.: изд. АН СССР, 1963. Вып. 1.
С. 161-167.
Бианки В. В., Карпович В. Н., .Макаров В. В., Татаринкова Н. Н.
Экология обыкновенной гаги во внегнездовый период;//Тр. Кандалакшского.
гос. заповедника: М., Лесная промышленность, 1967. Вып. V. С. 5-39.
Бианки В. В. Урожайность ягод водяники на островах Кандалакшского
залива и потребление их животными/./Продуктивность дикорастущих
ягодников и их oic. исполь;). - Кировск, 1972. С. 207-209.
Борисов А. А. Климаты СССР. - М.: Просвещение, 1967. 296 с.
Бострем Г. Г. Внутренние воды//География Архангельской области.
7-8 класс. Архангельск, 1983. С. 30-37.
Бреслина И. И. Приморекие луга Кандалакшского залива Белого моря//Биол.-флорист.
Исслед. в связи с охраной природы в Заполярье. - Апатиты, 1980.
С. 132-143.
Бреслина И. И. Растения и водоплавающие птицы мореких островов Eieuneie
Субарктики. - Л.: Наука, 1987. 200 с.
Боч М. С., Мазинг В. В. Экосистемы болот СССР. - Л.: Наука, 1979.
187 с. Возжинская В. Б. Донные макрофиты Белого моря. - М.: Наука,
1986. 191 с. Войлошникова Н. А., Войлошников В. Д. Реки//Архангельская
область. Экон.-геогр. хар-ка. - Архангельск, 1967, 456 с.
Волкова Л. А., Драбкова В. Г., Летанская Г. И. и др. Гидробиологическая
характеристика//Лимнологический очерк Вялозера//Большие озера Кольского
п-ова - Л., Наука, 1975. С. 70-105.
Гасконский А.М. Растительность//Архангельская область. Экон.-геогр.
хар-ка. - Архангельск, 1967. 456 с.
Герд С. В. Кормовые ресурсы oзеp Карело-Финской ССР//Рыбное хозяйство
Кар.-Фин. ССР//Тр. I научн.-техн. конф. по рыбн. пром-ти Кар.-Фин.
ССР. Октябрь 1945 г. Петрозаводск, 1947. Вып. VI. С. 102-108.
Гибет Л. А. Опыт типологии водных охотничьих угодий северо-запада
Карело-Финской ССР и иаснрсдслснис водоплавающих irruu//Elo.'i.
Моск. о-ва испытат. нр111)одь1. - М., 1953. Т. VIII, вып. 5. с.
21-29.
Голиков А. Н., Бабков А. И., Голиков А. А. и др. Экосистемы Онежского
эалива и сопредельных участков Бассейна Белого моря//Экосист. Онежского
залива Белого моря. - Л.: Изд. ЗИН АН СССР, 1985. С. 20-87.
Грибова С. А. Кольские кустарниковые и кустарничковые тундры. Южные
тундры. Предтундровые редколесья//Растительность Европ. части СССР.
- Л.: Наука. 1980. С. 62-69, 83.86.
Григорьев С. В. Некоторые закономерности озерности Карелии//VII
сессия уч. сов. по пробл. "Биол. ресурсы Белого моря и внутр.
водоемов Карелии". Март 1968 г.//Тез. докл. - Петрозаводск,
1968. С. 101-102.
Григорьев С. В., Грицевская Г. Л. Каталог озер Карелии. - М.-Л.:
Изд. АН СССР,1959, 240 с.
Грицевская Г. Л. Реки Карельского побережья Белого моря//Сессия
уч. сов. по пробл. "Теорет. основы рац. использ., воспроизвед,
и повышен, рыбных и нерыбных ресурсов Белого моря и внутр. водоемов
Карелии". 12-15 марта 1963 г. -Петрозаводск, 1963, С. 9-10.
Гурьянова Е. Ф. Условия жизни в Белом море//Рыбн. хоз-во Кар-фин.
ССР//Тр. I научн.-техн. конф. по рыбн. пром-ти Кар.-Фин. ССР. Октябрь
1945 г. -Петрозаводск, 1947. Вып. VI. С. 61-73.
Гурьянова Е. Ф. Краткие результаты гидробиологических исследований
Мезенского залива летом 1952 г.//Мат. по комплекс, изуч. Белого
моря. - М.-Л.: Изд. АН СССР. 1957. Кн. 1. С. 252-281.
Гурьянова Е., Закс И., Ушаков П. Литораль Кольского залива. Ч. III.
Условия существования на литорали Кольского залива//Тр. Ленингр.
о-ва естествоиспыт. -Л., 1930. Т. X. Вып. 2, С. 17-101.
Давыдов Л. К. Гидрография СССР (воды суши). Часть II. Гидрография
районов. - Л.: Изд. ЛГУ, 1955. 600 с.
Данилевский Н. Я. Рыбные и звериные промыслы на Белом и Ледовитом
Морях//Исслед. о сост. рыбол. в России. - Спб., 1862. Т. 6. 257
с.
Добровольский А. Д., Залогин Б. С. Моря СССР (природа, хозяйство).
М.: Мысль, 1965. 352 с. :,
Добровольский А. Д., Залогин Б. С. Моря СССР. - М.: Изд. МГУ, 1982.
192 с.
Дороганевская Е. А. Химизм плодов дикорастущих ягодников и их лекарственное
значснис//Продукт. дикорастущих ягодников и их хоз. использ. -Киров,
1972. С. 38-38.
Зенкевич Л. А. Биология морей СССР. М.: Изд. АН СССР, 1963. 740
с.
Зенкевич Л. А. Моря СССР, их флора и фауна: М.: Учпедгиз, 1956.
424 с.
Зубакин Г. К., Дементьев А. А. О современных тенденциях климата
Белого моря//Прир. и хоз-во Севера. - Мурманск, 1987. Вып. 15. С.
27-34.
Иванов А. И. Каталог птиц СССР. Л.: Наука, 1976. 276 с.
Иванова С. С. Зоопланктон губы Чупа//Maт. по комплекс, изуч. Белого
моря. -М. - Л.: Изд. АН СССР, 1963. Ч. II С. 17-31.
Извекова Э. И., Качанова-Львова А. А., Соколова Ю. Ю. Фауна озер
острова Великого и полуострова Киндо Кандалакшского залива Белого
моря//VII сессия уч. cов. по пробл. "Биол. ресурсы Белого моря
и внутр. водоемов Карелии". Март 1968 г//Тез. докл. Петрозаводск,
1968. С. 101-102.
Исаченко Т. И. Североевропейские еловые леса//Растит, европ. части
СССР. -Л Наука, 1980. С. 82-83.
Исаченко Т. И., Лавренко Е. И. Ботанико-географическое районирование
//Растит европ. части СССР. - Л.: Наука, 1980. С. 10-20.
Казакова О. Н. Ландшафтное районирование Мурманской области//Северо-запад
европ. части СССР. - Л.: Изд. ЛГУ, 1972. Вып. 8. С. 134-157.
Кальберг 3. А. Геологическое строение, полезные ископаемые и рельеф//Архангельская
область. Экон. геогр. хар-ка. - Архангельск, 1967. 456 с.
Калякина Н. М- Биотопы беломорской Литорали//Пробл. науч., рац.
использ. и охраны природ, ресурсов Белого моря//Тез. докл. III регион,
конф. Кандалакша, 1987. Кн. I С. 168-170.
Камшилов М. М. Некоторые новые данные о зоопланктоне Белого моря//Мат.
по комплекс, изуч. Белого моря. - М.-Л.: Изд. АН СССР, 1957. С.
305-314.
Карпенко А. С. Сосновые леса//Растит. европ. части СССР. - Л.: Наука,
1980. С. 117-123.
Китаев С. 11. Экологические основы биопродуктивности озер разных
природных cii. - М.: Наука, 1984. 207 с.
Крючков В. В. Север на грани тысячелетии. - М.: Мысль, 1987. 270
с.
Кузнецов В. В. Белое море и биологические особенности его флоры
и фауны. -М.-Л.: изд. АН СССР, 1960. 322 с.
Купецкий В. Н. Стационарные полыньи в Белом море//Вопр. гидр. сев.
Mopей//Tp. ГОИН. - Л.: ГИМИЗ, 1961. Вып. 64. С. 78-92.
Лесков А. И. Геоботанический очерк приморских лугов Малоземельского
побережья Баренцева моря. - Ботанический журнал, 1936. Т. 21. №
1. С. 95-116.
Липатова В. В. Растительность пойм//Растит. европ. части СССР. -
Л.: Наука, 1980. С. 346-372.
Лобза Н. Г. Гидрохимические условия в районе Нюхчи и в Сорокской
губе в летний период//Мат. по комплекс. изуч. Белого моря. - М.-Л.:
изд. АН СССР, 1963. Ч. II. С. 5-16.
Лоция Белого моря. Картфабрика ВМФ, 1957. 336 с
Луканин В. В., Ошурков В. В., Бергер В. Я. Особенности распределения
и запасы мидии в Кандалакшском и Онежском заливах Белого моря//Повыш.
продукт, и рац. использ. биол. ресурсов Белого моря//Мат. I координ.
совещ. - Л.: БАН СССР, 1982. С. 156-157.
Максимова М. II. Сравнительная гидрохимическая характеристика рек
Терского, Кандалакшского, Поморекого и Лямицкого побережий Белого
моря//Ссссия уч. сов. по пробл. "Теорет. основы paц. использ.,
воспроизв. и :повыш. рыбных и нерыбных ресурсов Белого моря и внутр.
водоемов Карелии". 12-15 марта 1963 г. - Петрозаводск, 1963.
С. 7-9.
Моллюски Белого моря. - Л.: Наука, 1987. 325 с. ;
Мухомедияров Ф. В. Биология и промысел второстепенных промысловых
рыб Карельского побережья//Мат. по комплекс. изуч. Белого моря.
- М.-Л.: изд. АН СССР, 1963-б. Ч II С. 131-143..
Мухомедияров Ф. В. Ихтиофауна губы Чупа//Мат. по комплекс, изуч.
Белого моря. - М.-Л.; изд. АН СССР, 1963-а. Ч. II. С. 90-99.
Наумов А. Д., Бабков А. И., Луканин В. В., Федяков В. В. Гидрологическая
и биоценотическая характеристика Мезенского залива Белого моря//Экол.
исслед. донных организмов Белого моря. - Л.: 3ИH АН СССР, 1986.
С. 64-90.
Наумов А. Д., Луканин В. В., Федяков В. В. Количественное распределение
фауны и флоры Двинского залива Белого моря//Повыш. продукт, и рац.
использ. биол. ресурсов Белого моря//Мат. I координ. совещ. - Л.:
БАН СССР, 1982. С. 66-67.
Наумов А. Д., Луканин В. В., Федяков В. В. Донные сообщества Бассейна
Белого моря в районе Терского берега//Пробл. изуч., рац. использ.
и охраны природ, ресурсов Белого моря//Тез. докл. III регион, конф.
- Кандалакша, -.1987. Кн. II С. 204-206.
Наумов А. Д., Федяков В. В. Введение//Моллюски Белого моря. - Л.:
Наука, 1987. С. 6-13.
Невесский К. И., Медведев В. С., Калиненко В. В. Белое море. Седиментогенез
и история развития в голоцене. -М.: Наука, 1977. 236 с.
Никитина О. Н. Характеристика донной фауны озера Мураканского//Пробл.
изуч., paц. использ. и охраны природ, ресурсов Белого моря//Тез.
докл. регион. конф. - Архангельск, 1985. С. 150-151.
Нинбург Е. А. Экологическая структура мелководных губ Белого моря//Пробл.
изуч. рац. использ. и охраны природ, ресурсов Белого моря//Тез.
докл. регион. конф. - Архангельск, 1985. С. 151-153.
Паленичко 3. Г. Жизнь Белого моря. - Петрозаводск, 1968. 148 с.
Паракецов И. А. Экология некоторых донных рыб прибрежной зоны Белого
моря//Сессия уч. сов. по пробл. "Теорет. основы рац. использ.,
воспроизвод. и повыш. рыбных и нерыбных ресурсов Белого моря и внутр.
водоемов Карелии". -Петрозаводск, 1963. С. 35-36.
Перцова Н. М. Зоопланктон Кандалакшского залива Белого моря//Биол.
Белого моря: Тр. Беломор. биол. станции. - М.: Изд. МГУ, 1970. Т.
3. С. 34-35.
Покровский В. В., Новиков Н. И. Озера Карелии и их рыбохозяйственное
значениа//0зера Карелии. Природа, рыбы и рыбное хозяйство. Справочник
-Петрозаводск, 1959. С. 9-85.
Прони ?? М. В. Экология прибрежных донных сообществ Мурманского
побережья Баренцева моря. - Л.: Наука, 1971. 128 с.
Прыгункова Р. В., Перцова Н. М., Слонова С. А. и др. Зоопланктон
и зообентос как часть экосистемы Белого моря и кормовая база рыб//Пробл.
изуч., рац. использ. и охраны природ, ресурсов Белого моря//Тез.
докл. регион, конф. -Архангельск, 1985. С. 17-23.
Раменская .М. Л. Анализ флоры Мурманской области и Карелии. - Л.:
Наука, 1983. 216 с.
Раменская М. Л., Андреева В. II. Определитель высших растений Мурманской
области и Карелии. - Л.: Наука, 1982. 432 с.
Рихтер Г. Д., Чикишев А. Г. Север европейской части СССР. - М.:
Мысль, 1966. 238 с.
Романов А. А. О климате Карелии. - Петрозаводск, 1961. 140 с.
Рыкова С. 10. Численность и биотопическос распределение птиц в Пинежском
заповеднике//Размещ. и числ. животных на Европейском Севере// Коми
фил. АН СССР. - Сыктывкар, 1986. № 74. С. 46-55.
Салазкин А. А. О развитии зоопланктона в малых озерах северо-запада//У111
сессия уч. сов. по пробл. "Биол. ресурсы Белого моря и внутр.
водоемов Европ. Севера". Ноябрь 1969 г. - Петрозаводск, 1969.
С. 38-40.
Севастьянов Г. Н. Размещение тетеревиных в зависимости от урожая
ягодников в Коми АССР//Плодовитость дикорастущих ягодников и их
хоз. испольа. - Киров, 1972. С. 203-205.
Скарлато О. А., Алексеев А. 11. Пути повышения биологической продуктивности
Белого моря//Итоги и перспект. изуч. биол. ресурсов Белого моря.
- Л.: 3ИH АН СССР, 1983. С. 5-13.
Советский энциклопедический словарь. 2-е изд. - М.: 1982. 1560 с.
Соколов В. Е. Систематика млекопитающих. - М.: Высшая школа, 1973.
432 с.; 1977. 496 с.; 1979. 528 с.
Сорокин А. Л., Дмитриенко А. И. Донные ландшафты южного прибрежья
Баренцева моря//Вопр. геогр. океана.//Teз. докл. I Всес. конф).
Калининград, сентябрь 1983 г. - Л., 1983. С. 73-74.
Сорокин А. Л., Касабов Р. В., Ванюхин Б. И. Комплексные исследования
подводных ландшафтов прибрежных районов Белого моря//Повыш. продукт,
и рац. использ. биол. ресурсов Белого моря//Мат. I координ. совещ.
- Л.: БАН СССР, 1982. С. 22-23.
Тимонов В. В. Главные особенности гидрологического режима Белого
моря//Сб. памяти Ю. М. Шокальского. - M.-.Л. изд. АН СССР, 1950.
Т. 2. С. 206-235.
Турков В. Г., Шишкин Н. А. Опыт составления таблиц средней многолетней
продуктивности дикорастущих ягодников на Европейском Севере (Вологодская,
Архангельская области и Коми АССР)//Продукт. дикорастущих ягодников
и их хоз. использование, - Киров, 1972. С. 154-156.
Уралов Н. С. Некоторые особенности сезонной и многолетней изменчивости
положения внешней границы льдов Баренцева моря//Вопр. гидрол. сев.
Mopей//Tp. ГОИН. - Л.: ГИМИ3, 1961. Вьш. 64. С. 39-77.
Федоров Н. И. Влияние экологических условий на химический состав
плодов рябины обыкновенной//Продукт. дикорастущих ягодников и их
хоэ. иснольз. -Киров, 1972. С. 33-36.
Федяков В. В. Особенности распределения моллюсков Двинского залива
Белого моря//Повыш. продукт, и paц. использ. биол. ресурсов Белого
моря. Мат. I координ. совещ. - Л.: БАН СССР, 1982. С. 88-89.
Цветков В. Ф., Нечаев В. Н., Казаков Л. А. Леса на краю Земли. -
Мурманск, 1983. 160 с.
Цветков В. Ф., Чертовский В. Г. Классификационные типологические
схемы лесов и лесорастительное районирование Мурманской области.
- Архангельск, 1978. 36 с.
Цинзерлинг Ю. Д. Материалы по растительности северо-востока Кольского
полуострова//Тр. сов. по изуч. природ, ресурсов. Сер. Кольская.
- М.-Л.: изд. АН СССР, 1935. Вып. 10. С. 5-150.
Цинзерлинг Ю. Д., Косинская Е. К. Материалы к характеристике пресноводной
растительности северо-востока Кольского нолуострова//Тр. сов. но
илуч. природ, ресурсов. Сер. Кольская. - 'М.-Л.: Над. АН СССР, 1935.
Вып. 10. С. 151-152.
Чернов Е. Г. Что растет на Кольской земле//Природа Мурманской области.
-Мурманск, 1964. С. 115-135.
Черепанов С. К. Сосудистые растения СССР. - Л.: Наука, 1981, 510
с.
Шкляревич Г. А. Зообентос малых лагун Порьей губы Кандалакшского
залива Белого моря//Гидробиол. исслед. в заповедниках СССР. Teз.
докл. Всес. совещ. 17-21 апреля 1989 г. (Борок Ярославской обл.)
- М., 1989. С. 135-136.
Юрковская Т. К. Травяно-лишайниково-моховые болота. Травяно-сфагново-гипновые
болота//Растит. европ. части СССР. - Л.: Наука, 1980. С. 328-338.
Яковлев Ф. С., Воронова В. С. Типы лесов Карелии и их природное
районирование. - Петрозаводск, 1959. 191 с.
Burger A. E., Simpson М. Diving depths of Atlantic Puffins and Common
Murres. Auk, 1986, 103, № 4, р. 828-830.
Piatt J. F., Nettleship D. N. Diving depths от four alcids. Auk,
1985, 102, № 2, р. 293-297. Наверх
|